
שאלון מערכת: שעורי בית מולקולרית 4
מולקולרית 4 שאלה 1
באדם ישנם 3 קודוני סיום (stop codon) ידועים. מי מהבאים אינו קודון עצירה באדם?
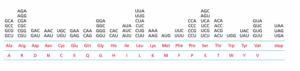
תשובה א‘. הקוד הגנטי (Genetic code) מתאר את הקומבינציה של 3 נוקלאוטידים ב-mRNA אשר כל אחד מכונה קודון (Codon). כל קודון מקודד לח. אמינו מסוימת. ישנו קודון אחד שמשמש כקודון התחלה (Start codon) שהוא AUG. קודון זה מקודד לח. האמינו מתיונין (Methionine) הן באאוקריוטים והן בפרוקריוטים, כאשר באחרונים מחוברת אליו קבוצת פורמיל (Formyl). וכן ישנם 3 קודונים אשר לא מקודדים לח. אמינו אלא לחלבון שנקרא פקטור שחרור (Release factor) שתפקידו להפסיק את קידוד ה-mRNA, קודונים אלו הם UAA, UAG ו-UGA. אני אוהב לזכור אותם לפי המשפט: “או-אה”,”או-אג” ו-“עו-גה”.
UGG מקודד לחומצת אמינו טריפטופן ומכיוון שזהו הקוד היחיד לחומצה אמינו זו מומלץ לזכור
להרחבה – Alberts, מהדורה שביעית, עמ’ 374-375 (Stop codons) ועמוד 359 (Genetic code).
מולקולרית 4 שאלה 2
מי מהבאים הינו תוצר ביניים בשלב הסינתזה של AA-tRNA-His בתהליך התרגום?
תשובה ד‘. תהליך טעינת ה-tRNA בחומצת האמינו המתאימה לו ולאנטי-קודון (Anti-codon) שלו, נעשית ע”י האנזים Aminoacyl tRNA synthetase. בשלב הראשון, האנזים מאקטב את החומצה האמינית ע”י AMP מ-ATP, תוך שחרור פירופוספט (Pyro-phosphate) – פעולה זו נקראת Adenylylation שבה נוצר ה-Adenylated amino acid. בשלב הבא, האנרגיה האגורה בקשר אמינואציל-AMP משמשת להעברת החומצה האמינית ל-tRNA המתאים, כך שמקבלים אמינואציל-tRNA (Aminoacyl-tRNA) –פעולה זו נקראת Esterification מכיוון שנוצר פה קשר אסתרי (Ester bond) שבו ישנו קשר בין קבוצת הקרבוקסיל של ח. האמינו לפחמן של קבוצת הריבוז (Ribose) של אחד הנוקלאוטידים ב-Aminoacyl-tRNA. חשוב לשים לב כי אנזימים אלו נחלקים ל-2 קבוצות עיקריות: Class I אשר מחברים את ח. האמינו לפחמן 2′ בריבוז, ו-Class II אשר מחברים אותה לפחמן 3′ בריבוז. בכל מקרה ח. אמינו תמיד תגיע לעמדה 3′ בריבוז עקב ריאקציית הטרנס-אספריפיקציה (Transesterification). לכל סוג של ח. אמינו קיים אנזים אשר אחראי להטענתו (ולכן יש כ-20 אנזימים כאלו). לסיכום, תוצר הביניים מתייחס לריאקציה הראשונה של תהליך הטענת ה-tRNA שבה מאקטבים את ח. האמינו (היסטדין במקרה הזה) ויוצרים AMP-His ולכן זו התשובה הנכונה.
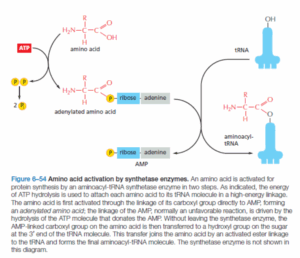
שלילת מסיחים:
א’. במולקולה AMP-tRNA-His ישנם שלושה מרכיבים: AMP, tRNA וח. אמינו היסטדין (His). לא יכול להיות מצב שבו נראה את ה-tRNA מחובר לח. אמינו כלשהי יחד עם ה-AMP כי ה-AMP יושב על המקום שבו הם צריכים להתחבר. צריך לזכור שהקשר ביניהם מתרחש בזכות ה-AMP אשר עזיבתו מאפשרת את החיבור בין הקרבוקסיל של ח. אמינו לריבוז של ה-tRNA. ולכן מסיח זה אינו נכון.
ב’. כפי שהוסבר מעל, אין תוצר ביניים שכולל בתוכו פוספט איאורגני – Pi. וכמו כן, חובה לזכור שה-ATP וה-AMP שנוצר מפירוקו משמשים ליצירת הקשר עתיר האנרגיה שמחבר בין ח. אמינו ל-tRNA ואין סיבה שפוספט כלשהו ימשיך להיות מחובר למולקולה כאשר הקשר כבר נוצר.
ג’. הפירו-פוספט אינו מתחבר בכלל לתוצרי הביניים. ריאקציית הפירוק של ATP ל-AMP ושיחרור ה-PPi יוצרת את הח. אמינו הטעונה ב-AMP. כמו כן הפירו-פוספט מפורק מיד לאחר מכן ל-2 פוספטים חופשיים.
להרחבה – Alberts, מהדורה שביעית, ע”מ 361-364 (tRNA charge).
מולקולרית 4 שאלה 3
איזה מוטציה תוביל לחלבון ארוך במיוחד?
תשובה ב‘. קודוני הסיום (Stop codons) הם השלושת הנוקלאוטידים שעליהם מתחבר פקטור השחרור (Release factor) שגורם לניתוק הפוליפפטיד מהריבוזום ולסיום תהליך התרגום (Translation termination). ה-Release factors הם חלבונים שמסוגלים להתחבר לאחד מקודוני הסיום – UAA, UAG ו-UGA (אני אוהב לזכור אותם כ”או-אה”,”או-אג” ו-“עו-גה”), כאשר הם נמצאים ב-A-site של הריבוזום – האתר שעליו נכנסים ה-tRNA שטעונות בח. האמינו החדשות בכל פעם. אם יש מוטציות ובעיות שונות שפוגעות בקודון הסיום ה-Release factor לא יוכל להתחבר לריבוזום ולכן תהליך התרגום ימשיך ונקבל פוליפפטיד ארוך מהרגיל. בהקשר השאלה, כאשר תתרחש מוטציה באנטי-קודון (Anti-codon) של ה-tRNA אשר תגרום להחלפת ציטוזין (Cytosine) לאורציל (Uracil) – ריאקציה שנקראת דה-אמינציה (Deamination), האנטי-קודון UCA מסוגל כעת להתחבר לקודון הסיום UGA. יכולתו של ה-tRNA שטעון בטריפטופן (Trp) להיקשר לקודון הסיום יגרום לריבוזום להמשיך לסנתז (ישים טריפטופן במקום להפסיק את התרגום) ולכן נקבל חלבון ארוך מהרגיל.
הערת המחבר – ישנו מקרה ספציפי שבו מסוגלת מולקולת tRNA מיוחדת שטעונה בח. אמינו סלנוציסטאין (Selenocysteine) להתחבר לקודון טרמינציה (UGA), לא ידוע למה דווקא קודון ספציפי פעם משמש לסיום ופעם אחרת משמש לח. האמינו סלנוציסטאין וכיצד הריבוזום יודע להבדיל ביניהם, אך חשוב להכיר מקרה זה.
שלילת מסיחים:
א’. אין דבר כזה tRNAstop. הפקטור שמהווה כגורם העוצר את התרגום נקרא Release factor ואינו סוג של tRNA מכיוון שהוא אינו עשוי מ-rRNA אלא מח. אמינו. מה שכן, הוא אכן דומה במבנה המרחבי שלו ל-tRNA ולכן הוא מסוגל להיכנס ל-A-site של הריבוזום ולגרום לטרמינציית התרגום.
ג’. חובה להיות ערניים ולא להתבלבל, השאלה מדברת על תהליך התרגום פגום שבו נוצר חלבון ארוך מהרגיל. ולכן מוטציה בתהליך השעתוק תפגע ביצירת גדיל ה-mRNA (ותיצור גדיל ארוך מהרגיל) ולא תגרום ליצירת חלבון ארוך מהרגיל מכיוון שקודון הסיום של התרגום עדיין תקין (מכיוון שרצף הטרמינציה של השעתוק נמצא באזור הלא-מקודד של ה-mRNA – נקרא UTR (ה-Untranslated region).
ד’. הפקטור eIF-2 הוא פקטור אינציאציה של תהליך התרגום באאוקריוטים אשר מחובר ל-GTP ומתחבר ל-Initiator tRNA–methionine (בקיצור Met–tRNAi) אשר מחובר ל-P-site תת-היחידה הקטנה בריבוזום. כאשר ה-GTP שעל eIF2 מפורק ל-GDP ו-Pi הוא מתנתק מתת-היחידה הקטנה ומאפשר לתת-היחידה הגדולה להתחבר עליהלכדי יצירת הריבוזום השלם. כאשר תתרחש מוטציית Loss of function ב-eIF2 שלב האינציאציה יפגע ולכן לא תתרחש כלל יצירת הפוליפפטיד, ולכןמסיח זה אינו נכון.
להרחבה – Alberts, מהדורה שביעית, ע”מ 368-375 (Translation), ע”מ 375-376 (Selenocysteine), ע”מ 455 (UTR)
מולקולרית 4 שאלה 4
RNA סינטטי המכיל את הרצף הבא …AUAAUAAUAAU… והודגר אצל חיידקים. בהנחה ולא נדרש קודון התחלה לרצף, כמה פפטידים שונים יוכלו להיווצר ממנו?
תשובה ד‘. מסגרת הקריאה (Reading frame) היא רצף הנוקלאוטידים (Nucleotides) שב-mRNA שממנו יקודדו את החלבון. כזכור, מסגרת הקריאה בנויה מקודונים (Codons) שהם רצף של שלושה נוקלאוטידים צמודים שמהווים כ”קוד” לח. אמינו מסוימת. באופן גורף ניתן להגיד שלכל mRNA, יש שלוש מסגרות קריאה אפשריות, אך התלות בקודון ההתחלה היא זו שמכתיבה את מסגרת הקריאה הנכונה עבור ה-mRNA הנתון. בכללי, ניתן ליצור שלושה צירופים מכל רצף של mRNA. פירוט: (1) כאשר הקודון הראשון מתחיל מהנוקלאוטיד הראשון. (2) כאשר הקודון הראשון מתחיל מהנוקלאוטיד השני. ו-(3) כאשר הקודון מתחיל מהנוקלאוטיד השלישי (ראו איור בע”מ 334). בהקשר השאלה, מהרצף …AUAAUAAUAAU… ניתן ליצור שלושה מסגרות קריאה: (1) רצף של AUA-AUA-AUA (הקודון הראשון מתחיל מהנוקלאוטיד הראשון).(2) רצף של UAA-UAA-UAA (הקודון הראשון מתחיל מהנוקלאוטיד השני), ו-(3) AAU-AAU-AAU (הקודון הראשון מתחיל מהנוקלאוטיד השני). כאשר הרצף השני של הקודונים UAA מהווה כרצף של קודוני סיום (Stop codons) ולכן הרצף לא יקודד לפפטיד בכלל. ולכן נוכל לקבל רק כ-2 פפטידים מהרצף הנתון.
הערת המחבר – חשוב להכיר ולזכור בעל פה את שלושת קודוני הסיום – UAA, UAG ו-UGA (“או-אה”, “או-אג” ו-“עו-גה”).
להרחבה– Alberts, מהדורה שביעית, ע”מ 358-359 (Reading frame) וע”מ 374-375 (Stop codons).
מולקולרית 4 שאלה 5
מי מבין המשפטים הבאים הקשורים לסינתזת חלבונים נכון?
במה השאלה עוסקת: Translation.
תשובה ב‘. הפקטור Elongation factor Tu (בקיצור EF-Tu) הוא פקטור אלונגציה של תהליך התרגום (Translation) בפרוקריוטים. פקטור זה, מביא את ה-Aminoacyl-tRNA לעמדה A (A-site) בריבוזום (Ribosome). כדי לבצע את תפקידו, החלבון חייב לקשור GTP, ובכדי להתנתק מהקומפלקס עם סיום התפקיד(הכנסת ה-tRNA לריבוזום), החלבון עושה הידרוליזה ל-GTP באמצעות פעילות GTPase אינטינזית, ובכך משתחרר מהקומפלקס. פקטור זה עוד יחד עם הפקטור EF-G אשר יחד פועלים בכל סיבוב (Cycle) של הכנסת ח. אמינו אחת לפוליפפטיד. האחרון אחראי לתנועת הטרנסלוקציה (Translocation) של הריבוזום לעבר הקודון הבא (גם הוא משתמש ב-GTP). הפקטורים הללו נקראים EF1 ו-EF2 באאוקריוטים, בהתאמה. ה-EF-Tu תורם גם חלק משמעותי ליכולת ה-Proofreading של הריבוזום, ע”י כך שהוא משהה את פירוק ה-GTP ולכן נשאר מחובר ל-tRNA על מנת לבדוק שיש התאמה מלאה של האנטי-קודון (Anti-codon) שב-tRNA לקודון שב-mRNA.
שלילת מסיחים:
א’. פעילות של האנזים פפטידיל טרנספראז (Peptidyl transferase) מבוצעת על ידי אלמנט ה-RNA שבריבוזום, ולא על ידי האלמנט החלבוני.כלומר, מדובר כאן בפעילות של Ribozyme – רנ”א בעל פעילות קטליטית. אנזים זה אכן משתתף בפעילות של סינתזת החלבונים (תרגום) מכיוון שהוא יושב ב-23S שנמצאת בתת-היחידה הגדולה (50S) של הריבוזום הפרוקריוטי (וב-28S באאוקריוטים), אך הוא איננו חלבון ולכן מסיח זה אינו נכון.
הערת המחבר – ישנם 2 אנטיביוטיקות חשובות שהכרחי להכיר בהקשר של האנזים: (1) כלורמפניקול (Chloramphenicol) שמעכבת אותו בפרוקריוטים. ו-(2) אניסומיצין (Anisomycin) אשר מעכבת אותו באאוקריוטים.
ג’. פקטור שחרור (Release factor) הוא לא tRNA. אומנם שניהם דומים במבנה המרחבי שלהם, ושניהם מסוגלים להיכנס ל-A-site של הריבוזום, אך למעשה הפקטור שחרור בנוי מחלבון לעומת ה-tRNA שהוא סוג של גדיל RNA. אין tRNA אשר מסוגל להתחבר לקודון הסיום (Stop codon) ולגרום להפסקת התרגום.
הערת המחבר – ישנו מקרה ספציפי שבו מסוגלת מולקולת tRNA מיוחדת שטעונה בח. אמינו סלנוציסטאין (Selenocysteine) להתחבר לקודון טרמינציה (UGA), לא ידוע למה דווקא קודון ספציפי פעם משמש לסיום ופעם אחרת משמש לח. האמינו סלנוציסטאין וכיצד הריבוזום יודע להבדיל ביניהם, אך חשוב להכיר מקרה זה.
ד’. כפי שנאמר מעל, הפקטור EF-Tu פועל באלונגציה ולא בשלב האינציאציה (Initiation) ולכן מסיח זה איננו נכון.
להרחבה – Alberts, מהדורה שביעית, ע”מ 368-369 (EF-Tu), ע”מ 377 (Selenocysteine),ע”מ 368 (Peptidyl transferase) וע”מ 374-375 (Release factor).
מולקולרית 4 שאלה 6
התגלתה מוטציה אשר הופכת קודון של חומצת אמינו לקודון סיום. מה מהבאים נכון לגבי מוטציה זו?
תשובה א‘. מוטציית פסק (Nonsense) היא סוג של מוטציה נקודתית (Point mutation) אשר בה נוקלאוטיד אחד משתנה לאחר, וגורםעקב כך לשינוי הקודון (Codon) שמקודד לח. אמינו מסוימת להיות קודון סיום(Stop codon). במקרה כזה היווצר במהלך התרגום (Translation) פוליפפטיד יותר קצר. במצב הנורמלי שבתא,ישנו מנגנון שנקרא Nonsense-mediateddecay (בקיצור NMD) אשר אחראית לאיתור וגילוי גדילי mRNA אלו והשמדתם. תהליך זה מתרחש בזכותקומפלקסים מיוחדים שנקראים Exonjunction complexes (בקיצור EJC) אשר נמצאים על ה-mRNA באתר החיתוך (Splice site) של כל אינטרון (מתחברים לכל אתר לאחר השחבור). כאשר הריבוזום הראשון מתרגם את ה-mRNA הוא “מפיל” את כל ה-EJC שנמצאים על הגדיל (מכיוון שרץ לכל אורכו עד לאקסון האחרון שמכיל את קודון הסיום). כאשר קודון הסיום ימצא באמצע ה-mRNA, עקב מוטציית Nonsense שיצרה אותו, הריבוזום הראשון שיתרגם את הגן ייעצר באמצע (בקודון הסיום הלא מקורי) ולכן הוא לא יפיל את כל ה-EJC (ב-mRNA תקין לא אמורים להיות כלל EJC עליו). מכיוון שה-EJC נשארו על ה-mRNA יגיעו חלבונים מיוחדים שנקראים Upf אשר יצרו קומפלקס גדול שישלח את ה-mRNA לדגרדציה (Degradation). מנגנון ה-NMD חשוב הן לניצול אנרגטי יעיל למניעת בזבוזאנרגטי והן למניעת יצירת חלבונים מוטנטים שיכולים לגרום נזק לתא.
שלילת מסיחים:
ב’. המושג פולימורפיזם (Polymorphism) מתאר שינוי שחל בגנום שהוא לא נדיר, כלומר שינוי בגנום שהוא נפוץ יחסית. כאשר מדובר על שינוי שבו מוחלף נוקלאוטיד אחד זה יקרא SNP (שזה Single-nucleotide polymorphisms) וכאשר השינוי מתרחש עקב הכנסה או מחיקה (Insertion or Deletion) של מספר נוקלאוטידים זה יקרא אינדל (Indel), במקרה של שינוי קטן, ו-CNV (שזה Copy number variations) במקרה של שינוי נרחב. השינויים הללו אינם גורמים ליצירת חלבון קצר יותר מהרגיל, אלו הבדלים בין גנומים שונים אצל אוכלוסיות שונות, לרוב שונות זאת היא שקטה ואינה באה לידי ביטוי בפנוטיפ כלשהו.
ג’. מוטציית סלף (Missense) היא מוטציה שבה יש שינוי בנוקלאוטיד אחד המשנה את הקודון לקבלת חומצה אמינית שונה. מוטציה זו יכולה להיות שמרנית (Conservative) – שינוי לחומצה אמינית בעלת אותן התכונות (א-פולרית\פולרית\טעונה או ארומטית), או לא-שמרנית (Non-conservative) – שינוי לחומצה אמינית בעלת תכונות שונותמאלו של המקורית. דוגמה נפוצה ניתן למצוא במחלת האנמיה החרמשית (Sic – שינוי מגלוטמט (טעונה שלילית) לואלין (אפולרית)-מוטציה המביאה לאנמיה חרמשית. בשני המצבים נקבל פפטיד השונה בחומצה אמינית אחת.
ד’. מוטציה שקטה (Silence mutation) היאמוטציה שבה יש שינוי בנוקלאוטיד אשר לא מביא לשינוי בחומצה אמינית. למעט מתיונין (Methionine) וטריפטופן (Tryptophan), לכל שאר החומצות האמינו יש בין 2-6 קודונים, מה שמקנה את האפשרות ששינוי בנוקלאוטיד אחד (בדרך כלל השלישי בקודון) לאיביא לשינוי בחומצה האמינית, ועל כן יתקבל חלבון זהה לזה שהיה לפני המוטציה. גם מוטציה זו אינה הופכת לקודון סיום ולכן מסיח זה אינו נכון.
להרחבה – Alberts, מהדורה שביעית, ע”מ 374-375 (Stop codons), ע”מ 378-379 (Nonsense-mediated mRNA decay), ע”מ 324, 525-526 (Polymorphisms), ע”מ 253-254 (Silent mutation) ועמוד 358 (Genetic code).
מולקולרית 4 שאלה 7
איזה מהקודונים הבאים יזוהה על ידי האנטיקודון UAC?
תשובה ד‘. ה-Wobble effect או Wobble base-pair היא תופעה שבה הצימוד בסיסים (Base pair) בין הנוקלאוטיד השלישי בקודון (Codon) לבין הנוקלטוטיד הראשון באנטי קודון (Anti-codon), לא מתקיים רק לפי ווטסון וקריק. הקשר בעמדה זו יכול להיות מתירני ורופף יותר, ומאפשר צימוד בין זוגות לא קונבנציונלים של נוקלאוטידים. כאשר ניגשים לפתור שאלה כזו, חשוב לזכור שקריאת הקודונים היא משמאל לימין, כאשר השמאלי ביותר יהיה קצה 5′ והנוקלאוטיד בקצה הימיני ביותר יש קצה 3′. בנוסף, חשוב לשים לב שהאנטי-קודון (שהיא אחת הזרועות של ה-tRNA) מתחברת ל-Codon ב-mRNA בצורה של מהופכת של Anti-parallel– כלומר קצה 5′ מתחבר לקצה 3′. תופעת Wobble מאפשרת לקודון השלישי (בקצה 3′ של ה-mRNA) להתחבר לנוקלאוטידים שונים באנטי-קודון, ניתן לראות בתמונה את האפשרויות הקיימות:
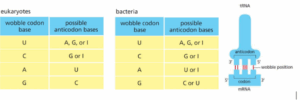
כפי שניתן לראות, בתשובה ד’ יש G בקצה 5′ שמסוגל להתחבר ל-C בקצה ה-3′ של האנטי-קודון, ו-G בעמדה 3′ שמסוגל להתחבר ל-U בקצה 5′ של ה-tRNA (=וובל). ולכן תשובה זו הנכונה.
שלילת מסיחים:
א’. כזכור, הקודון AUG הוא הקודון היחיד שמסוגל לקודד לח. האמינו מתיונין (Methionine), אשר משמש בנוסף כקודון ההתחלה (Start codon) שאליו התחבר ה-Initiator tRNA המיוחד – בפרוקריוטים זה יהיה מתיונין שמחובר אליו קבוצת פורמיל (Formyl-methionine). ה-C שבקצה 3′ של האנטי-קודון אינו מסוגל להתחבר ל-G של הקודון בקצה 5′ ולכן מסיח זה אינו נכון.
ב’. גם במקרה זה ה-C שבקצה 3′ של האנטי-קודון אינו מסוגל להתחבר ל-G של הקודון בקצה 5′ ולכן מסיח זה אינו נכון. חשוב לא להתבלבל בין קצה 3′ לקצה 5′.
ג’. אינוסין (Inosine) הוא נוקלאוטיד מיוחד אשר נוצר כאשר הבסיס החנקני Hypoxanthine מתחבר לבסיס הסוכרי ריבוז (Ribose). האינוסין יכול להיווצר עקב תהליכי דה-אמינציה (Deamination) של אדנין (Adenine) ספונטניים או ע”י אנזים מיוחד שנקרא A-to-I editing enzyme (בקיצור ADAR). את האינוסין רואים בעיקר במולקולות ה-tRNA (ניתן לזכור זאת עקב הבסיס הריבוזי שלו ולאהדיאוקסי-ריבוז שבדנ”א).
להרחבה – Alberts, מהדורה שביעית, ע”מ 360 (Wobble), ע”מ 358-360, ע”מ 291-292 (Hypoxanthine), ע”מ 337 (Inosine) וע”מ 451 (ADAR).
מולקולרית 4 שאלה 8
בוצעה אנליזה גנטית עבור רצפו של גן ספציפי בפרט החולה במחלה תורשתית. החוקר זיהה מוטציה נקודתית- שינוי של נוקלאוטיד C לנוקלאוטיד T שגרמה להחלפת חומצת האמינו גלוטמין לקודון פסק. כל ההיגדים הבאים נכונים למעט:
תשובה ד‘. בניתוח גנטי (Genetic analysis) הכוונה היא לכל שיטה גנטית שבה ניתן לבחוןולאמוד את הגנום כגון Polymerasechain reaction (בקיצור PCR), קריוטיפ (Karyotype), ריצוף גנטי (DNA sequencing) וכדומה. כאשר התרחשה מוטציה נקודתית (Point mutation), כלומר החלפה של נוקלאוטיד אחד באחר אשרגורם לשינוי ח. אמינו של אותו הקודון (Codon) לקודון סיום (Stop codon) הכוונה היא Nonsense mutation. מוטציית Nonsense יוצרת חלבון קצר יותר מהרגיל עקב הופעתו שלקודון סיום באמצע האזור המקודד ב-mRNA. לרוב, מוטציה זו מתוקנת ע”י מנגנון ה-Nonsense-mediated decay (בקיצור NMD) שמסוגל לזהות את הקודון הנ”ל ע”יחלבונים מיוחדים שנקראים Exon junction complexes(בקיצור EJC) שנוכחותם מעידה על תרגום לא מלא של הגדיל –הריבוזום (Ribosome) הראשון שמתרגם את ה-mRNA אמור ל”הפיל” (להוריד) את כל ה-EJC מה-mRNA. החלפה של נולקאוטיד אחד באחר איננה מוסיפה או גורעת מסך מספר הנוקלאוטידים ולכן לא תתרחש מוטציית Frameshift שבה יש הזזה של מסגרת הקריאה ושינוי כל הקודונים עקב זאת, ולכן מסיח זה אינו נכון ואותו נסמן.
שלילת מסיחים:
א’. התמרה פשוטה (Transition mutation) היא החלפה של נוקלאוטיד אחד בנוקלאוטיד אחר כאשר הנוקלאוטידים המוחלפים הם מאותו הסוג, כלומר כאשר פורין (Purine) מוחלף בפורין או כאשר פירמידין (Pyrimidine) מוחלף בפירמידין אחר. בהשאלה זו ההחלפה הייתה בין ציטוזין לתימין ששניהם מסוג פירימידין ולכן מסיח זה הגיוני ונכון.
הערת המחבר – כזכור,פורין הוא סוג של בסיס חנקני (Nitrogenbase)שמורכב מ-2 טבעות חנקניות כגון Adenine, Guanine ו-Hypoxanthine. פירימידין הוא גם סוג של בסיס חנקני אשר מורכב מטבעת חנקנית אחת בלבד, כגון Cytosine, Thymine ו-Uracil.
ב’. פולימורפיזם (Polymorphism) הוא מושג המתאר שינוי נפוץ שחל בגנום באוכ’ מסויימת. כאשר מדובר על שינוי שבו מוחלף נוקלאוטיד אחד זה יקרא Single-nucleotide polymorphisms (בקיצור SNP) וכאשר השינוי מתרחש עקב הכנסה או מחיקה (Insertion or Deletion) של מספר נוקלאוטידים זה יקרא אינדל (Indel), במקרה של שינוי קטן, ו-CNV (שזה Copy number variations) במקרה של שינוי נרחב. לרוב, שינויים פולימופיים אלו בגנום אינם גורמים לפנוטיפ כלשהו או למחלה מסוימת. הפולימורפיזם זהבעצם השונות שיש בין בני אדם בתכונות שונות כגון צבע עור, גובה וכדומה. מכיוון שברוב המקרים לא תהיה משמעות קלינית ככל הנראה לא מדובר במוטציה הנקודתית שיוצרתקודון סיום שלו יש השלכות משמעותיות על תפקוד החלבון והתא.
ג’. מוטציה נקודתית היא החלפה שלבסיס חנקני אחד באחר, ולכן מסיח זה נכון (כלומר לא נסמן אותו).
להרחבה– Alberts, מהדורה שביעית, ע”מ 253-254 ועמוד 358(Point mutation), ע”מ 378-379 (Nonsense-mediated mRNA decay), ע”מ 525-526 (Polymorphisms)
מולקולרית 4 שאלה 9
מסגרת הקריאה (open reading frame) של אנזים מסוים מורכבת מ102 נוקלאוטידים. מה תהיה, ככל הנראה, ההשפעה של מוטציית השמטה נקודתית של הנוקלאוטיד ה-73?
תשובה ד‘. תחילה נחשב כמה חומצות אמינו מכיל הפפטיד לפני השפעת המוטציה. נחסיר מ-102 שלוש, שכן, השלשה האחרונה מקודדת לקודון סיום ולא לחומצה אמינית: 102-3=99. את 99 נחלק בשלוש- 99\3=33, כלומר, החלבון המקורי, מכיל 33 חומצות אמינו. מוטציית השמטה (Deletion mutation) מביאה להזזת מסגרת הקריאה (Frameshift) מנקודת המוטציה ו-Downstream אליה (כך שכבר ניתן לפסול את מסיח ב’).עכשיו ניתן לענות על השאלה באופן אינטואיטיבי. אם החלבון מכיל 33 חומצות אמינו בסה”כ, והמוטציה התרחשה בנוקלאוטיד ה-73 מתוך 102, רוב החלבון יהיה לפי הרצף התקין, כך שניתן לשלול את סעיף א’ ואת סעיף ג’. ולאלה שאוהבים בכל זאת לחשב, אם המוטציה התרחשה בנוקלאוטיד ה-73, אזי, שעד הנוקלאוטיד ה-72, הכל תקין. 72נוקלאוטידים הינם 24 חומצות אמינו (72\3) – אם מתוך 34 חומצות אמינו, 24 נשארות תקינות, זה משאיר אותנו עם 10 קודונים שהשתנו במורד הזרם למוטציה, קודון אחד מתוך העשרה זהו קודון סיום שלא מקודד לחומצה אמינית, ולכן נקבל שינו ב-9 החומצות האמיניות האחרונות – 34-24=10. 10-1=9. חשוב לשים לב, שגם אם רוב רצף החלבון תקין, אין זה אומר שהחלבון יהיה פונקציונלי באופן מלא. המוטציה יכולה לפגוע במבנהו המרחבי ולגרום לירידה בתפקודו. בנוסף. מוטציה המשנה את מסגרת הקריאה, פעמים רבות מביאה גם להופעת קודון סיום מוקדם, שכן הסיכוי להופעתו הוא 3\64.
להרחבה– Alberts, מהדורה שביעית, ע”מ 358-360 (Reading frame) וע”מ 374-375 (Stop codons).
מולקולרית 4 שאלה 10
לפניך תגובות המתרחשות במהלך הטענת tRNA, אותן מזרז האנזים אמינו-אציל-tRNA-סינתטאז. איזה מהמסיחים מתאר נכונה את הקשר בין התגובות?
- amino acid + ATP-> aminoacyl-AMP+PPi
- amino acid + ATP+ tRNA-> aminoacyl-tRNA+AMP+PPi
- amino acyl-AMP + tRNA-> aminoacyl-tRNA+AMP
תשובה ב‘. בשביל לענות על השאלה, עלינו להכיר את שני שלבי ראקציית ההטענה שמתבצעים על ידי האנזים אמינואציל tRNA סינתתאז (Aminoacyl tRNA synthetase). בשלב הראשון, האנזים מאקטב את החומצה האמינית ע”י AMP מ-ATP, תוך שיחרור פירופוספט (Pyro-phosphate). בשלב הבא, האנרגיה האגורה בקשר אמינואציל-AMP משמשת להעברת החומצה האמינית ל-tRNA המתאים, כך שמקבלים אמינואציל-tRNA. באנזים קיימים 3 אתרים במרכזיים:
1) אתר קישור לח. האמינו.
2) אתר קישור ל-AMP.
3) אתר קישור ל-tRNA.
בראקציה המלאה משתמשים בחומצה אמינית אחת, tRNA מתאים ובמולקולת ATP אחת, ששוות ערך לשתי מולקולות אנרגיה, עקב שיחרורו של הפירופוספט לתמיסה. בנוסף לפירופוספט, משתחרר בסופו של דבר גם ה-AMP ששימש לאיקטוב החומצה האמינית, ומקבלים tRNA טעון שישמש בבניית פפטידים חדשים בתא.
להרחבה– Alberts, מהדורה שביעית, ע”מ 361-363 (tRNA charge).
מולקולרית 4 שאלה 11
מה לא נכון בנוגע לריבוזום?
תשובה ג’. הריבוזום (Ribosome) הוא קומפלקס מיוחד אשר מבצע את פעולת התרגום (Translation) של גדילי ה-mRNA בכלל האורגניזמים השונים. קומפלקס זה הוא סוג של ריבונוקלאופרוטאין אשר מורכב הן מחלבונים והן מחומצות גרעין אשר נחלק ל-2 תתי-יחידות עיקריות: גדולה וקטנה. באאוקריוטים הגדולה נקראת 60S (לפי מקדם השקיעה שלה – Sedimentation coefficient) והקטנה 40S. וכן בפרוקריוטים יש את ה-50S וה-30S, בהתאמה. לריבוזום יש 3 אתרים (Sites) עיקריים שמאפשרים את תהליך התרגום. הראשון נקרא A-site ואליו נכנס ה-Aminoacyl-tRNA (ומכאן שמו), השני P-site שבו מתרחש פעולת יצירת הקשר הפפטידי (ע”י האנזים Peptidyl transferase שנמצא בתת-היחידה 28S או 23S) כלומר באתר זה תהיה המולקולה Peptidyl-tRNA, ובאתר האחרון שנקרא E-site ה-tRNA הריק יוצא (Exit). תתי-היחידות של הריבוזום אכן מקודדות ע”י אותו ה-RNA polymerase בפרוקריוטים (מכיוון שקיים רק רנ”א פולימראז אחד בהם שמסוגל לקודד את כל סוגיה-RNA), אך באאוקריוטים, ישנם מספר RNA polymerases שמשתתפות בייצורו של הריבוזום, דבר ראשון ה-RNA polymerase II אשר אחראי לייצור החלבונים שלו וכן ה-RNA polymerase I שאחראי לייצור הרוב ה-rRNA שבונים אותו. בנוסף, גם הרנ”א פולימראז 3 תורם חלק קטן מהריבוזום ע”י ייצור ה-5S rRNA שמהווה כחלק מתת-היחידה הגדולה (60S), ולכן מסיח זה אינו נכון ואותו נסמן.
שלילת מסיחים:
א’. כפי שנאמר מעל, אכן ה-40S וה-30S הן תתי-היחידות הקטנות שלא האאוקריוטים והפרוקריוטים, בהתאמה. כמו כן, חשוב להכיר שה-40S מורכבת מ-rRNA של 18S וה-30S מ-16S (שמהווה כרצף התחברות ל-Shine–Dalgarno sequence).
ב’. לריבוזום יש כ-4 אתרי קשירהלגדילי RNA, ישנם את שלושת האתרים העיקריים שתוארו מעל:ה-A-site, P-site וה-E-site (שקושרים tRNA) ויש את אתר הקשירה ל-mRNA שנקרא מן הסתם mRNA binding site.
ד’. נכון. תת-היחידה הקטנה מתחברת ראשונה לגדיל ה-mRNA בשלב האינציאציה של התרגום (Translation initiation).
להרחבה– Alberts, מהדורה שביעית, עמ’ 365-368 (Ribosome) ועמ’ 369-371 (Translation).
מולקולרית 4 שאלה 12
השתקת גנים באמצעות RNAi מתרחשת לרוב על ידי _______ של גן המטרה
תשובה ב‘. מנגנון ה-RNAi (RNA interference) הוא צורה רגולטורית של התא על מנת לבקר את ביטוי הגנים שבו. במנגנון זה משתתפים שלושה סוגי RNA עיקריים: (1) miRNA (microRNA). (2) siRNA (small interfering RNA) ו-(3) piRNA (piwi-interacting RNA). ה-miRNA הן המולקולות הכי שכיחות במנגנון זה אשר מבצעות רגולציה על כ-1/3 מהגנים המקודדים לחלבונים. הן עושות זאת ע”י עיכוב או פירוק ה-mRNA בציטופלזמה כאשר מתחברות לקומפלקס RISC (RNA-induced silencing complex). מולקולות ה-siRNA משמשות כמנגנון הגנהכנגד גנומים זרים החודרים לתא (כמו של ווירוסים או חיידקים). הן מיוצרות ע”י האנזים Dicer שבתגובה ל-dsRNA ארוך וזר בתא הוא חותך אותו ויוצר דו-גדילים קצרים (נקראים siRNA) שמתחברים לאנזים RISC שמפרק גדילים תואמים ל-siRNA שחובר אליו, כך הם מפרקים את שאר גדילי הגנום הזר שנמצאים בתא. האחרון, piRNA, פועל בצורה מעט שונה ופועל כמנגנוןהגנה רק בתאי נבט (Stem cells) כנגד טרנספוזציה (Transposition). בשונה מהשניים הקודמים כאן האנזים Dicer אינו מעורב, אך המנגנוןהפירוק של יצירת זיווג בסיסים עם גדיל המטרה (גדיל ה-RNA של הטרנספוזון) נשארזהה. כפי שנאמר בהתחלה, המנגנון הכי שכיח ב-RNAi הוא ה-miRNA אשר מונעות תרגום (Translation).
שלילת מסיחים:
א’. מנגנון הפרעתה-RNA (RNAi) אכן יכול למנוע שעתוק (Transcripation), וזאת ע”י יכולתן של מולקולות ה-siRNA ליצור מבני הטרוכרומטין (Heterochromatin) בכרומוזומים. בפעולה זו, מעורב האנזים Dicer שמתחבר ל-Argonaute ועוד מספר חלבונים על מנת ליצור את הקומפלקס RITS (RNA-induced transcriptional). ה-RITS מסוגל לגייס חלבוניםרבים אשר מבצעים מודיפיקציות רבות על ההיסטונים (Histones) ולגרום להם לעלות את רמת הדחיסהשל הכרומוזום. RITS מבצע זאת ע”י התחברות לגדיל שמשועתק ויוצא מה-RNA polymerase II בזמן הסנתוז. תהליך זה נעשה בעיקר סביב הצנטרומר (Centromere). כמובן שתהליך זה לא שכיח כמו הפעילות של ה-miRNA ולכן מסיח זה אינונכון.
ג’. מניעת תהליך השחבור (Splicing), שהוא תהליך הוצאת האינטרונים (Introns) מגדיל ה-mRNA והשארת האקסונים (Exons) בגדיל, איננה חלק ממנגנון הפרעתה-RNA.
ד’. תהליך הפוליאדנילציה (יצירת זנב ה-Poly-A ב-mRNA) הוא תהליך חשוב מאוד שמקנה ל-mRNA הגנה מפני אקסונוקלאזות (Exonucleases), מאפשר את הוצאת הגדיל מהגרעין (Transport) ואת התחלת התרגום שלו (Translation initiation). אומנם תהליך זה מאוד חשוב ומהווה כמרכיב קריטי מביטוי הגן אך הואאיננו קשור למנגנון ה-RNAi.
להרחבה – Alberts, מהדורה שביעית, ע”מ 462-468 (RNAi), ע”מ 341-348 (Splicing) וע”מ 348-349 (Polyadenylation).
מולקולרית 4 שאלה 13
לאיזה אזור בtRNA נקשרת החומצה האמינית?
תשובה ד‘. מולקולת Transfer RNA (בקיצור tRNA) היא סוג של RNA שמקודד ע”י RNA polymerase III אשר משמש כסוג של מתאמת (Adaptor) בין ה-mRNA לח. האמינו. ישנם מעל ל-20 סוגי tRNA (אחת לכל ח. אמינו) כל אחת באורך ממוצע של80 נוקלאוטידים. ה-tRNA עובר מספר רב של מודיפיקציות בתהליך שנקרא “עיבוד רנ”א” (RNA editing) שבמהלכו מוסיפים ומורידים ממנו נוקלאוטידים מסוימים ואף משנים אותם לגמרי – בממוצע 10 אחוז מה-tRNA עובר מודיפיקציות ושינויים. במבנה הראשוני (Primary structure) נראה את רצף הנוקלאוטידים שמרכיבים את הגדיל, במבנה השניוני (Secondary structure) הרצף מקבל צורה שדומה לעלה תלתן (Cloverleaf) שבה ניתן לראות את הזרועות (Arms) השונים של ה-tRNA כגון את זרוע האנטי-קודון (Anti-codon arm), ה-D-loop (מבנה של Stem & loop שמכיל מספר נוקלאוטידים מסוג Dihydrouridine=D), ה-Acceptor stem (שהיא הזרוע בקצה 3′ שמתחברת לח. אמינו המתאימה) וכו’. לבסוף, במבנה השלישוני (Tertiary structure) נראה את המבנה התלת-ממדי שנראה כאות “L” הפוכה. כפי שנאמר, ה-Acceptor stem (ה”זרוע המקבלת”) היא הקצה 3′ של ה-tRNA שמסתיימת ברצף הנוקלאוטידים CCA, כאשר אל ה-A מתחברת ח. אמינו בקשר קוולנטי חזק. ולכן תשובה זו הנכונה.
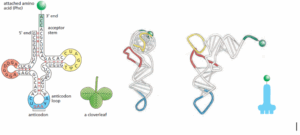
שלילת מסיחים:
א’. ה-D loop הוא חלק שבנוי בצורת Stem & loop ב-tRNA אשר מכיל מספר נוקלאוטידים מסוג Dihydrouridine. לולאה זו אינה מתחברת לח. אמינו ולכן מסיח זה אינו נכון.
ב’. כמובן שח. אמינו לא מתחברת לאנטי-קודון, כפי ששמו מעיד עליו, הוא ה”אנטי” של הקודון שנמצא ב-mRNA ואמור להשלים אותו ולהתחבר לאליו ולכן אינו יכול להתחבר בנוסף גם לח. אמינו, ולכן מסיח זה אינו נכון.
ג’. כפי שנאמר מעל, ח.אמינו מתחברת לרצף CCA אשר נמצא בקצה 3′ ב-tRNA, חשוב לזכור את הכיווניות ולא להתבלבל ביניהם.
להרחבה– Alberts, מהדורה שביעית, ע”מ 359-361 (tRNA).
מולקולרית 4 שאלה 14
מי מהבאים נקשר ישירות לCAP?
תשובה ב‘. המודיפיקציה הראשונה שנעשית עלה-mRNA היא יצירת ה-5′ Cap שלו. מודיפיקציה זו נעשית כאשר ה-RNA polymerase II סנתז כבר כ-25 בסיסים בערך וזנב ה-CTD (ה-C-terminal domain) שלו מזורחן בעמדת ה-Ser5 מה שמאפשר לשלושת אנזימי ה-Capping ליצור את מבנה זה (Phosphatase, Guanyl transferase ו-Methyl transferase). ישנם שלושה פקטורי אינציאציה (Initiation factors) שחשוב להכיר בהקשר תרגום (Translation) באאוקריוטים:
1) eIF2 – אחראי לקישור האמינואציל-tRNA ההתחלתי (Initiator tRNA) לריבוזום. פקטור זה דורש GTP לפעולתו שעובר הידרוליזה כשהפקטור מתנתק מהריבוזום. פעולה אשר תורמת ליכולת ה-Proofreading של הריבוזום.
2) eIF4E – פקטור שנקשר ישירות ל-5′-Cap, אליו מתחבר הפקטור eIF4G.
3) eIF4G – חלבון פיגום (Scaffold protein), הנקשר מצד אחד ל-eIF4E, ומצד שני לחלבונים הקשורים לזנב הפולי A בקצה 3′ ב-mRNA שנקראים PABP. ע”י כך פקטור זה גורם להתעגלות המולקולה, ולקירוב שתי הקצוות שלה, פעולה המייעלת את התרגום באופן מרחבי (תתי היחידות של הריבוזום מתנתקות בקצה 3′ שנמצא קרוב לקצה 5′ שממנו יתחיל סיבוב תרגום חדש).
שלילת מסיחים:
א’. מסיח שבה לבלבל בין תהליכי השעתוק (Transcription) והתרגום, וכן בין ה-RNA polymerase לריבוזום. חשוב להיות מרוכזים ולא ליפול במסיחים הללו.
ג’. ה-40S היא תת היחידה הקטנה של הריבוזום האאוקריוטי. תת-יחידה זו מתחברת לקודון ההתחלה (Start codon) שהוא ה-AUG הראשון ב-90% מהמקרים, אך בחלק מהמקרים הוא מסוגל להתחיל ב-AUG מאוחר יותר (Downstream). ה-40S אינו מתחבר ישירות ל-5′-Cap אלא לפקטור eIF4E שמחובר אליו, ולכן מסיח זה אינו נכון.
ד’. ה-Poly-A-binding protein (בקיצור PABP) הם חלבונים מיוחדים שמתחברים לאורכו של זנב ה-Poly-A אשר מתחברים אליו בזמן שה-Poly-A polymerase (בקיצור PAP) מסנתז אותו. ה-PABP חשובים מאוד לשמירה כנגד אנדונוקלאזות (Endonucleases), העברה מהגרעין לציטופלזמה ולאינציאציית התרגום.
להרחבה– Alberts, מהדורה שביעית, ע”מ 337-338 (5′-Cap), ע”מ 348-349 (PABP), ע”מ 373-374 (Translation initiation) ועמוד 366 (Ribosome subunits).
מולקולרית 4 שאלה 15
איזה שלב אינו תואם את הריאקציה המתרחשת בו?
תשובה ד‘. בשלב הטרמינציה (Termination) של התרגום יתחבר חלבון מיוחד שנקרא פקטור שחרור (Release factor) כאשר יופיע אחד מקודוני הסיום (Stopcodons)באתר A של הריבוזום. כזכור, קודוני הסיום הם UAA, UAG ו-UGA (אני אוהב לזכור אותם כ”או-אה”, “או-אג” ו-“עו-גה”) והם מסמנים לפקטור זה להיכנס לריבוזום ולגרום לריבוזים (Ribozyme) שנקרא פפטידיל-טרנספראז (Peptidyl transferase) לקטלז הכנסת מולקולת מים (H2O) לקצה ה-C של הפוליפפטיד מה שיגרום לניתוקו מה-tRNA ומהריבוזום בכלל. לאחר מכן הריבוזום יתנתק מה-mRNA ויפורק לשתי תתי-היחידות שלו בסדרת ריאקציות שדורשות GTP ולכן מסיח זה אינו נכון ואותו נסמן.
שלילת מסיחים:
א’. ישנו מקרה מיוחד שבו נמצא קודון עצירה UGA בתוך רצף ה-mRNA שאינו נחשב לקודון עצירה אלא כקודון מיוחד שאליו תתחבר tRNA שנושאת עליה ח. אמינו מסוג סלנוציסטאין (Selenocysteine). בשביל שריאקציה זו תתרחש צריך להיות מבנה מיוחד של Stem &loop ב-3’UTR שיסמן לפקטור תרגום מיוחד (שנקרא Selenocysteine-specific translation factor) לגייס את ה-tRNA הנ”ל.
ב’. פירוק ה-GTP ל-GDPב-eIF2 מתרחש כאשר ה-Initiator tRNA נמצא מול קודון ההתחלה AUG ב-mRNA וזה גורם לגיוס תת-היחידה הגדולה (60S) וליצירת הריבוזום השלם.
ג’. לרוב, מתיונין תמיד נקשרת לקודון הפתיחה בשלב האינציאציה, אך חשוב לזכור שח. אמינו זו יכולה להימצא גם באמצע הפוליפפטיד ולכן הכנסתה יכולה להתרחש גם בשלב האלונגציה באתר A (כזכור, בשלב האינציאציה ה-tRNA עם המתיונין נכנסת ישירות לאתר P בריבוזום – המקרה היחיד שזה קורה).
להרחבה – Alberts, מהדורה שביעית, עמ’ 368-375 (Translation).
מולקולרית 4 שאלה 16
כבר במהלך התרגום נצמדים חלבונים מיוחדים שנקראים שפרונים לפוליפפטיד המתהווה. איזה מהבאים אינו נחשב שפרון?
תשובה א‘. בכללי, השפרונים (Chaperones) הם חלבונים אשר עוזרים בקיפול רוב החלבונים בתא. חלבונים אלו, מתחברים לאזורים ההידרופוביים של הפוליפפטיד ומתחברים אליו על מנת שיתקפל בצורה האופטימלית שלו. ישנם סוגים רבים של שפרונים כגון Heat shock proteins (בקיצור Hsp), שפרונין ואף שפרונים שמתמחים בחיבור היסטונים לקומפלקס הנוקלאוזום (אשר נקראים Histone chaperones). ה-Upf הוא חלבון שלא קשור לקיפול חלבונים, חלבון זה מהווה כחלק ממנגנון ה- nonsense-mediated mRNA decay (בקיצור NMD). כזכור, מנגנון זה אחראי על גילוי ואיתור גדילי mRNA בעלי קודון עצירה מוקדם (Premature termination codon), וזאת בעזרת התחברות ה-Upf לקומפלקסים מיוחדים שנקראים Exon junction complexes (בקיצור EJC). לאחר כל שלב בתהליך השחבור (Splicing) מתחבר EJC אחד באתר החיתוך, וכאשר הריבוזום מגיע לתרגם את ה-mRNA בפעם הראשונה (נקרא לעיתים Pioneer round או Test round) הוא “מפיל” בכל פעם EJC כאשר עובר דרכו. כאשר הריבוזום נתקל בקודון עצירה מוקדם (שלא אמור להיות שם) ישארו עוד EJC על ה-mRNA שלא ירדו (כי הריבוזום לא הגיע עליהם), מה שמסמן לתא שה-mRNA לא עבר תרגום מלא ונורמלי. כאן ה-Upf נכנסים לפעולה, ומתחברים ל-EJC הנותרים ושולחים את כל הגדיל לפירוק.
שלילת מסיחים:
ב’. השפרון Hsp60 מגיע ממשפחת ה-Heat shock proteins אשר מסונתזים בעיקר במצבים של טמפ’ גבוהה. שפרונים אלו פועלים בעזרת ATP ונמצאים בעיקר במיטוכונדריה. שפרונים אלו מתחברים לפוליפפטיד במהלך התרגום וע”י כך מונעים קיפול לא נכון שלו.
ג’. החלבון Bip הוא סוג של שפרון ממשפחת ה-Hsp70 אשר עוזר לקפל חלבונים בתוך ה- Endoplasmic reticulum (בקיצור ER). כמו משפחת ה-Hsp60, גם שפרונים אלו מתחברים לאזורים ההידרופוביים של הפוליפפטיד ומשתמשים ב-ATP. כמו כן, הוא מקבלים עזרה מחלבונים נוספים שנקראים Hsp40 שמסייעים להם בהתחברות ובניתוק מהפוליפפטיד.
ד’. ה-TCP1 הוא שפרון שנמצא בציטוזול של אאוקריוטים. ולכן גם מסיח זה אינו נכון.
להרחבה– Alberts, מהדורה שביעית, עמ’ 378-379 (Upf), עמ’ 380-383 (Chaperones)
מולקולרית 4 שאלה 17
מה מהבאים לא נכון בהקשר של אפקט וובל?
תשובה ב‘. זיווג בסיסים מסוג Wobble הוא מקרה שבו האנטי-קודון (Anti-codon arm) של ה-tRNA מסוגל “לסבול” או “לקבל” חוסר התאמה כאשר מבצע זיווג בסיסים עם הקודון של ה-mRNA, וזה רק בעמדה הכי 5′ באנטי-קודון (שהיא גם העמדה הכי 3′ של הקודון). חוסר התאמה (Mismatch) זה, מאפשר לסוגי tRNA שונים, אשר דומים ב-2 מתוך 3 הבסיסים של האנטי-קודון והקודון, להתחבר לאותו הקודון גם אם הם שונים ואינם מתאימים בעמדה אחת מתוך האנטי-קודון. אפקט זה לא יכול להתרחש בעמדה 1 או 2 של האנטי-קודון אשר יותר “מחמירים” עם הדרישות להתאמה בזיווגי הבסיסים ביניהם. כמו כן, תופעה זו קיימת הן באאוקריוטים והן בפרוקריוטים, ולכן מסיח זה אינו נכון. ההבדלים בזיווגי הוובל שבין האאוקריוטים והפרוקריוטים כפי שניתן לראות בתמונה, נובעים מהמבנה השונה של הריבוזום ביניהם.
שלילת מסיחים:
א’. אכן עמדת הוובל (Wobble position) נמצאת בעמדה שנמצאת הכי במורד הזרם (Downstream), כלומר בקצה הכי 3′ של הגדיל, בקודון. וכךלהיפך בנוגע לאנטי-קודון, עמדה זו נמצאת הכי Upstream.
ג’. עצם זה שזיווג הבסיסים שבין האנטי-קודון לבין הקודון נעשה בידי 2 בסיסים בלבד ולא כל הבסיסים בקודון, מאפשר ל-tRNA שונים שמתאימים רק ב-2 בסיסים להתחבר לקודון. כלומר, ניתן להשתמש בפחות סוגים של tRNA על מנת להשלים את תהליך התרגום.
ד’. הנוקלאוטיד אינוזין נוצר בריאקציה של דה-אמינציה (Deamination) של אדנין (Adenine). בסיס זה יחסית נפוץ ב-tRNA, וכפי שניתן לראות, עקב הדמיון שלו לגואנוזין (Guanosine) הוא מסוגל להתחבר כמעט לכל שאר הבסיסים מלבד הגואנוזין, ולכן משפט זה נכון.
להרחבה – Alberts, מהדורה שביעית, עמ’ 359-361 (Wobble).
מולקולרית 4 שאלה 18
מי מהבאים לא מתאר נכונה את הפקטור שלו?
תשובה ג‘. הפקטור eIF4G הוא פקטור אאוקריוטי אשר פועל בשלב האינציאציה בתרגום (Translation initiation). פקטור זה נחשב כחלבון פיגום (Scaffold protein) אשר מהווה כ”גשר” בין קצה 3′ לבין קצה 5′ של ה-mRNA. כמובן שהוא לא מתחבר ישירות אל קצוות אלו, אלא דרך Poly-A binding proteins (בקיצור PABP) ו-eIF4E, בהתאמה. פקטור זה הכרחי ליצירת מבנה הפוליזום (Polysome) שנקרא גם פוליריבוזום (Polyribosome), שבו גדיל ה-mRNA מקבל צורה מעגלית שמאפשרת יצירת תהליך תרגום מתמשך עם מספר רב של ריבוזומים עליו בו-זמנית. אומנם, באאוקריוטים פקטור זה צריךאת ה-eIF4E על מנת שיוכל להתחיל את תהליך התרגום, אך בוירוסים יש מבנה מיוחד שנקרא Internal ribosome entry site (בקיצור IRES) שבו נוצר מבנה מיוחד של Stem & loop אשר מתחבר ישירות לפקטור eIF4G ללא צורך בפקטור eIF4E לרבות צורך ב-Cap על מנת להתחיל תרגום, ולכן מסיח זה אינו נכון.
שלילת מסיחים:
א’. אכן, הפקטור eIF4E מתחבר לתת-היחידה הקטנה ול-5′-Cap בתחילת התרגום. לאחר החיבור ביניהם, תת-היחידה הקטנה מתנתקת מהפקטור ומתחילה בתהליך הסריקה אחר קודון ההתחלה – AUG.
ב’. ה-eIF2B הוא סוג של Guanine nucleotide exchange factor אשר מאפשר את החידוש של הפקטור eIF2 ע”י הורדת ה-GDP ממנו. הורדה זו מאפשרת ל-GTP חדש להתחבר ל-eIF2 וכך לגרום לו להתחיל עוד סיבוב של תרגוםע”י קשירה מחודשת ל-Methionyl initiator tRNA הבא.
ד’. ה-eIF4F הוא גם פקטור אינציאציה חשוב אשר מעודד את קשירת תת-היחידה הקטנה של הריבוזום ל-AUG הקרוב ביותר לקצה 5′ של ה-mRNA ולכן פקטור זה אכן מפחית את תופעת ה-Leaky scanning שבו הריבוזום מדלג על ה-AUG הראשון (ולעיתים גם על הבא אחריו), מה שגורם ליצירת איזופורמים שונים של אותו החלבון (קצרים יותר מהרגיל).
הערת המחבר- למרות שעוד לא הגענו לחומר בסעיף ב’ עדיין אפשר לענות על השאלה דרך ידע שיש לנו מהמצגת הזו. זה מצב שעלול לקרות לפעמים במבחן הידע בו יש סעיף של שאלה שאנו לא מכירים. חשוב גם להתכונן למצב זה.
להרחבה – Alberts, מהדורה שביעית, עמ’ 368-376 (Translation factors) ועמ’ 457 (eIF4F+eIF2B).
מולקולרית 4 שאלה 19
מה מהבאים אינו נכון לגבי תהליך הRNA processing שמתרחש בtRNA?
תשובה ב‘. מולקולת ה-tRNA (שפירושה Transfer RNA) מקודדת ע”י RNA polymerase III באאוקריוטים, ולרוב מזוהה בשתי צורות עיקריות: הראשונה היא צורת התלתן (Cloverleaf) במבנה השניוני שלה (Secondary structure) והשנייה היא צורת האות “L” הפוכה במבנה השלישוני שלה (Tertiary structure). כמו הרבה מולקולות אחרות ה-tRNA עוברות גם עיבוד RNA על מנת להפוך לפעילים. בדומה לתהליך השחבור (Splicing) שעוברים גדילי ה-mRNA גם ה-tRNA עוברים סוג של שחבור אך לא ע”י הספלייסוזום בציטופלזמה, אלא מספר אנזימים אחרים שנמצאים בגרעין התא (Cell nucleus) כגון multifunctional tRNA ligase ואנדונוקלאז (Endonuclease) מיוחד. מכיוון שתהליך הספלייסינג נעשה בגרעין ולא בתא, תשובה זו אינה נכונה ונסמן אותה.
שלילת מסיחים:
א’. גדיל ה-tRNA נוצר כגדיל אחד ארוך אשר עובר חיתוך בקצותיו, תהליך זה נקרא Trimming (“קיטום”) והוא חשוב לקיפול נכון של המולקולה בהמשך.
ג’. אכן כל גדיל tRNA חייב לעבור מספר מודיפיקציות שונות, בממוצע אחד לכל 10 נוקלאוטידים יעבור מודיפיקציה או התמרה כלשהי. אחת ההתמרות הנפוצות ב-tRNA היא הדה-אמינציה (Deamination) של אדנין אשר יוצרת את הבסיס המיוחד אינוזין (Inosine).
ד’. כל העיבודים הנ”ל יכולים להתרחש הן בפרוקריוטים והן באאוקריוטים.
להרחבה – Alberts, מהדורה שביעית, עמ’ 359-361 (tRNA).
מולקולרית 4 שאלה 20
איזה מהמשפטים הבאים נכון לגבי shine Dalgarno sequence?
תשובה א‘. רצף השיין-דלגרנו הוא רצף קישור לריבוזום (Ribosome-binding site) אשר נמצא ב-mRNA הפרוקריוטי. ליתר דיוק, זהרצף הקישור לגדיל ה-16S rRNA שנמצא בתת-היחידה הקטנה של הריבוזום (30S). ה-16S יוצר זיווג בסיסים עם השיין דלגרנו, וכלל שמידת ההתאמה ביניהם (ככל שהרצף שיין-דלגרנו יהיה יותר קרוב לרצף הקונצנזוס – 5′-AGGAGGU-3′) תהיה יותר גבוהה כך הסיכוי שמסגרת הקריאה (ה-ORF) הזו תעבור תרגום (Translation) עולה. רצף זה נמצא תמיד במעלה הזרם (Upstream), כלומר יותר קרוב לקצה 5′, ביחס לקודון ההתחלה (AUG). בנוסף, חשוב להכיר כי ישנו תיווך של מספר פקטורי אינציאציה (Initiation factors) בין הרצף לריבוזום ושפעולת זיווג הבסיסים ביניהם תתרחש רק בנוכחותם.
שלילת מסיחים:
ב’. בניגוד לתרגום באאוקריוטים, שבו תת-היחידה הקטנה (40S) מתחברת ל-5′-Cap של ה-mRNA ומתחילה בתהליך סריקה (Scanning) של הגדיל עד שהיא מאתרת את קודון ההתחלה, בפרוקריוטים, תת-היחידה הקטנה של הריבוזום מתחברת ישירות לקודון ההתחלה ע”י קישור לרצף שיין-דלגרנו, ולכן מסיח זה אינו נכון.
ג’. כזכור, לפרוקריוטים יש תכונה חשובה שנקראת Polycistronic, שמתארת את יכולת ה-mRNA שלהם לקודד למספר חלבונים שונים, אך בניגודל תהליך השחבור החליפי (Alternative splicing) שגם יוצר חלבונים שונים באאוקריוטים, במקרה זה נקבל חלבון אשר שונה לגמרי ברצפו מחלבון אחר שגם קודד באותו ה-mRNA. ולכן באותו גדיל mRNA פרוקריוטי נראה מספר קודוני ההתחלה (AUG) מפוזרים ועקב כך מספר רצפי שיין-דלגרנו צמודים אליהם.
ד’. עקב חשיבותו הרבה של רצף זה, הואעובר הרבה בקרות בצורות שונות, למשל בקרה ע”י ריבוסוויץ (Riboswitch), רצף רנ”א רגיש טמפ’ (RNA Thermosensor), חלבונים מעכבי תרגום (Translation repressor protein) וכדומה.
להרחבה – Alberts, מהדורה שביעית, עמ’ 373-374 (Shine-dalgarno), עמ’ 365-367 (Ribosome subunits) ועמ’ 456-457 (Shine-dalgrano regulation).
מולקולרית 4 שאלה 21
השתקת גנים באמצעות RNAi מסוג miRNA מתרחשת על ידי _____ של גן המטרה
התשובה הנכונה היא ב’- מולקולות RNA interference מסוג miRNA מעכבות תרגום על ידי זיווג בסיסים עם mRNA המטרה. מולקולות ה- miRNA משועתקות על ידי RNA polymerase 2 ולאחר מכן עוברות תהליך עיבוד יחודי שכולל קישור לחלבונים ויצירת RNA induced silencing complex (RISC) שברגע שנוצר, מחפש את מולקולת mRNA המטרה על ידי זיווג ל- miRNA המעובד. אם יש זיווג בסיסים מושלם בין mRNA המטרה וה- miRNA אז mRNA המטרה ישלח לפירוק (נדיר ביונקים, נפוץ בצמחים. אם הזיווג בסיסים אינו מושלם, אז תרגום mRNA יעוכב, וה- mRNA ישלח לגופיפי P בציטופלסמה, שם כעבור זמן מסויים של עיכוב תרגום (מורחק מאזור ציטופלסמטי שכיח בריבוזומים) יעבור דגרדציה. פעילות miRNA היא בציטופלסמה, על כן,פעילותו היא במניעת תרגום ולא מניעת שעתוק, שחבור או פוליאדנילציה שבד”כ קורים בגרעין.
שלילת מסיחים:
א. כפי שהוסבר במסיח ב’, התפקיד של miRNA הוא בבקרת תרגום
ג. כפי שהוסבר במסיח ב’, התפקיד של miRNA הוא בבקרת תרגום
ד. כפי שהוסבר במסיח ב’, התפקיד של miRNA הוא בבקרת תרגום, בד”כ מניעת התרגום נעשית על ידי קיצור או ביקוע זנב ה- polyA ובכך ערעור יציבות ה- mRNA. אולם אין עצירה של תהליך הפוליאדנילציה.
להרחבה- Alberts עמודים 462-468
מולקולרית 4 שאלה 22
מה מהבאים מתאר נכונה את תופעת הleaky scanning?
תשובה ד‘. תופעת ה-Leaky scanning מתרחשת בשלב האינציאציה של תהליך התרגום (Translation initiation). כזכור בתחילת האינציאציה, תת-היחידה הקטנה של הריבוזום (40S), אשר מחוברים עליה ה-initiator tRNA והפקטור eIF2, מתחברת ל-5′-Cap של ה-mRNA ומתחילה בסריקה אחר קודון ההתחלה (AUG), כאשר קודון זה ימצא ויהיה בעמדה מול ה-P-site של הריבוזום (שבו נמצא ה-Initiator tRNA עם המתיונין עליו) ה- GTP שנמצא על ה-eIF2 יעבור הידרוליזה ויהפוך ל-GDP שיגרום לשיחרורו ולגיוס תת-היחידה הגדולה (60S) של הריבוזום לכדי יצירת הריבוזום השלם. אך לא תמיד ה-40S יזהה את ה-AUG הראשון שיתקבל בו בתהליך הסריקה, לעיתים תת-יחידה זו “תדלג” ותתחבר לקודון התחלה אחר שימצא יותר Downstream אליו. כאשר זה התרחש היווצר חלבון קצר יותר שיהווה כאיזופורם לחלבון המקורי והנורמלי שנוצר כאשר התרגום מתחיל מה-AUG הראשון. תופעה זו תלויה ברצפים שונים שנמצאים מסביב לקודון ההתחלה.
הערת המחבר – כזכור מביותא, בקצה N של הפוליפפטיד (N-termini) שנוצר לאחר התרגום יש רצף מיוחד שנקרא Signal sequence. רצף זה מהווה כרצף שמסמן לאן אותו חלבון מיועד ולאן ינוע בתא (או מחוצה לו), מכיוון שתהליך התרגום מתחיל בקצה ה-N טרמינלי של הפוליפפטיד וממשיך לשים ח. אמינו בכל פעם מחדש בקצה ה-C (קרבוקסילי) של הפוליפפטיד, תופעת ה-Leaky scanning משפיע ברוב הפעמים על ה-Signal sequence אשר שינוי בו יגרום לשינוי היעד של החלבון.
שלילת מסיחים:
א’. תהליך ה-Translation recoding הוא מקרה שבו הקידוד של mRNA מסוים מתרחש בצורה מעט שונה. למשל, במקרה שבו יש את קודון הסיום UGA (“עו-גה”) בתוך מסגרת הקריאה, אך מבנה מיוחד ב-3’UTR גורם לפקטור Selenocysteine-specific translation factor לגייס tRNA מיוחד של סלנוציסטאין (Selenocysteine) ולהמשיך בתרגום לאחר מכן. תהליך זה לא קשור לתהליך ה-Leaky scanning ולכן מסיח זה אינו נכון.
ב’. תהליך ה-Leaky scanning הוא תהליך שמתרחש רק באאוקריוטים, כיוון שתהליך הסריקה לא מתרחש בפרוקריוטים כי הריבוזום הפרוקריוטי מתחבר ישירות ל-AUG ע”י קישור לרצף ה-Shine-dalgrano שנמצא צמוד אליו, ולכן מסיח זה אינו נכון.
ג’. מנגנון ה-Nonsense-mediated mRNA decay (בקיצור NMD) הוא מנגנון אשר מתמודד עם קודון עצירה (Stop codon) שנמצא באמצע מסגרת הקריאה (ORF) של ה-mRNA. תהליך שמונע יצירה של חלבונים לא פעילים ומוטנטים ואינו קשור לתהליך ה”דילוג” של הריבוזום על קודוני ה-AUG.
להרחבה – Alberts, מהדורה שביעית, עמ’ 373-374 (Leaky scanning), עמ’ 378-379 (NMD), עמ’ 375-376 (Translation recoding)
מולקולרית 4 שאלה 23
מה הוא UTR?
תשובה ד‘. ה-UTR הם רצפים ב-mRNA אשר לא עוברים תרגום (Untranslated regions). ישנו רצף 5’UTR שנמצא בין ה-5′-Cap לבין האזור המקודד (Coding region) ויש 3’UTR שנמצא בין סוף האזור המקודד לזנב ה-Poly-A. לאזורים אלו תפקידים רבים, אחד מהם הוא רגולציית תרגום ה-mRNA, מכיוון שהם עשויים להכיל את הריבוסוויץ (Riboswitch), אתרי קישור ל-miRNA, אתרי IRES וכו’. מכיוון שאזורים אלו לאחר השעתוק (Transcription) עוברים שחבור (Splicing) ויוצאים מהגרעין הם נחשבים אקסונים (Exons) ולכן תשובה זו נכונה.
שלילת מסיחים:
א’. ה-Spliceosome הוא קומפלקס חלבון + RNA (מסוג snRNA) אשר אחראי על תהליך השחבור (Splicing). קומפלקס זה בנוי בעיקר מחלבונים ומ-5 סוגים של snRNA שנקראים U1, U2, U4, U5 ו-U6. כפי שהוסבר מעל, ה-UTR לא קשור לקומפלקס זה ולכן מסיח זה אינו נכון.
ב’. ה-UTR קיים הן בפרוקריוטים והן באאוקריוטים ולכן מסיח זה לא נכון.
ג’. כפי שנאמר מעל, ה-Open reading frame (בקיצור ORF) נתחם ע”י 2 ה-UTR וכמובן שהוא אינו עובר תרגום כלל.
להרחבה – Alberts, מהדורה שביעית, עמ’ 453-456 (UTR) ועמ’ 341-344 (Spliceosome).
מולקולרית 4 שאלה 24
מה הוא סדר הפעולות הנכון בתהליך הטענת הtRNA?
- tRNA מתחבר לAminoAcyl-tRNA synthetase
- שחרור tRNA
- ח. האמינו מתחברת לAminoAcyl tRNA-synthetase
- שחרור פירופוספט
- אקטיבציית ח. האמינו
- יצירת קשר אסטרי
- שחרור AMP
תשובה ב‘. האנזים Aminoacyl-tRNA synthetase אחראי על תהליך טעינת ה-tRNA (נקרא tRNA charging) בח. האמינו המתאימה לו. באאוקריוטים, ישנם כ-20 סוגים של Aminoacyl-tRNA synthetase בתאים, אחת לכל סוג של חץ אמינו, לעומתם, בפרוקריוטים יש פחות מ-20 מכיוון שכל Aminoacyl-tRNA synthetase מסוגל לטעון מספר סוגים של ח. אמינו. תהליך הטעינה: בהתחלה האנזים מתחבר לח. אמינו הנכונה, ואז הוא מבצע הידרוליזה ל-ATP ומשחרר פירו-פוספט לציטופלזמה. בתהליך זה הוא יוצר תוצר לוואי של Adenylated amino acid. לאחר מכן, האנזים מתחבר ל-tRNA המתאים ומשחרר את ה-AMP שהיה מחובר לח. אמינו. ע”י כך, נוצר קוולנטי חזק מסוג אסתר (Ester bond) בין ח. אמינו ל-tRNA. ולבסוף, האנזים משחרר את Aminoacyl-tRNA.
להרחבה – Alberts, מהדורה שביעית, עמ’ 361-363 (Aminoacyl-tRNA synthetase).
מולקולרית 4 שאלה 25
מי מהבאים נכון לגבי הטרנסלוקציה בתהליך התרגום?
תשובה ב‘. הריבוזום (Ribosome) בנוי חלבונים ו-rRNA אשר יוצרים 2 תתי-יחידות עיקריות – קטנה וגדולה. באאוקריוטים הם נקראים 40S ו-60S, בהתאמה, ובפרוקריוטים 30S ו-50S בהתאמה. במהלך שלב האלונגציה (Elongation) הריבוזום נע בכל פעם 3 נוקלאוטידים במורד הזרם (Downstream) ב-mRNA, תנועה זו נקראת טרנסלוקציה (Translocation). תהליך הטרנסלוקציה דורש פקטור אלונגציה שנקרא EF-G בפרוקריוטים ו-EF2 באאוקריוטים. פקטור זה מבצע הידרוליזה ל-GTP בכל מחזור לש טרנסלוקציה על מנת לגרום לשינוי קונפורמציה שיביא לתנועת תת-היחידה הגדולה קודון אחד קדימה ולאחר מכן לתנועת תת-היחידה הקטנה גם. בסיום תהליך זה הריבוזום זז בדיוק שלושה נוקלאוטידים במורד הזרם, מה שגורם לפתיחת ה-A-site בריבוזום ל-tRNA חדש.
הערת המחבר – אנטיביוטיקה חשובה שצריך להכיר בהקשר הטרנסלוקציה נקראת Cycloheximide. אנטיביוטיקה זו מעכבת את תנועת הריבוזום באאוקריוטים, ומשמשת רבות למחקר במעבדה.
להרחבה – Alberts, מהדורה שביעית, 365-369 ועמ’ 377 (Cycloheximide).
מולקולרית 4 שאלה 26
רצף האנטיקודון 5-UCG-3 יגיב עם איזה קודון?
- 5-CGA-3
- 5-CGG-3
- 5-GGC-3
- 5-AGC-3
תשובה ד‘. כדי לענות על שאלה זו, חשוב מאד להיות מסודרים מבחינת הכיוונים של הקודון והאנטיקודון.רצף האנטי קודון בשאלה – UCG יבצע צימוד בסיסים רגיל עם שני הנוקלאוטידים הראשונים שבקודון – CG. בשלב זה ניתן כבר לפסול מסיחים שלא מכילים את שני הנוקלאוטידים הראשונים האלה. עכשיו הגענו לעמדת ה-Wobble העמדה הראשונה באנטי-קודון והשלישית בקודון. U (יורודין) יכול לבצע צימוד בסיסים רגיל עם הנוקלאוטיד הקונבנציונלי שלו – A, ובנוסף יכול לבצע צימוד בסיסים עם הנוקלאוטיד G בפרוקריוטים בלבד. ולכן, קיימות שתי אופציות: CGA ו-CGG.
להרחבה – Alberts, מהדורה שביעית, ע”מ 358-361.
מולקולרית 4 שאלה 27
מה נכון לגבי שלב האלונגציה בתרגום חלבון?
תשובה ג‘. בשלב האיניציאציה, הגיעה המולקולה Aminoacyl-tRNA-Met לאתר P (P-site) בריבוזום. זוהי הפעם היחידה, שמתחילה מולקולה כזו בעמדה זו בריבוזום. מכאן ואילך, כל שאר המולקולות, יכנסו לקומפלקס האלונגציה דרך אתר A (A-site). כדי ליצור את הקשר הפפטידי בין הח. האמינו האחרונה שהוספה, לחומצה האמינית החדשה שהגיעה, תוקפת קבוצת הקרבוקסיל בח. האמינו הקודמת את הקבוצה האמינית בח. האמינו החדשה ליצירת קשר פפטידי. הריאקציה המרכזית הזאת מבוצעת ע”י האנזים פפטידיל טראנספראז (Peptidyl-transferase), שהוא חלק מתת היחידה הגדולה של הריבוזום (28S באאוקריוטים ו-23S בפרוקריוטים). פעולה זו חוזרת חלילה, עד ליצירת הפפטיד השלם, ומסתיימת כשהריבוזום מגיע לקודון הסיום (Stopcodon). לאחר כל הארכה בחומצה אמינית אחת, מתרחשת טרנסלוקציה (Translocation) של תת היחידה הגדולה ביחס לתת היחידה הקטנה, כך שאזורי ה-Acceptor stems של מולקולות ה-tRNA נמצאים באתרי P ו-E של תת היחידה הגדולה. בשלב הבא, סדרה של שינויים קונפורמטיביים מביאה לטרנסלוקציה של תת היחידה הקטנה. התזוזה מביאה אותה להחזיק את מולקולת ה-mRNA שלושה נוקלאוטידים Downstream, וגורמת ליציאה של ה-tRNA הריק מאתר E . כמו בשלב האינציאציה, גם כאן ישנם פקטורים העוזרים בראקציית האלונגציה. בפרוקריוטים אלו הפקטורים EF-G ו-EF-Tu ובאאוקריוטים אלו הפקטורים EF1 ו-EF2 (בהתאמה). כל ארבעת הפקטורים מנצלים מולקולת GTP ולארבעתם יש פעילות GTPase. בכל סיבוב אלונגציה יש שימוש בשני הפקטורים, ולכן שימוש בשתי מולקולות GTP.
שלילת מסיחים:
א’. הטרנסלוקציה (Translocation) שמתבצעת כל סיבוב בשלב האלונגציה גורמת לתזוזה של קודון שלם (שלושה בסיסים חנקניים=נוקלאוטידים) ולא של בסיס יחיד, ולכן מסיח זה אינו נכון.
ב’. כפי שנאמר מעל, בכל סיבוב אלונגציה יש שימוש בשני פקטורי האלונגציה, שכל אחד מהם עושה שימוש ב-GTP אחד, ולכן נעשה שימוש בשתי מולקולות GTP סה”כ בכל הארכה של חומצה אמינית אחת. ולכן מסיח זה אינו נכון.
ד’. בסיום כל שלב טרנסלוקציה אחד, הפפטיד המתהווה מחובר ל-tRNA שנמצא באתר P (אני אוהב לזכור זאת כמו “חניה” = “Parking site”) דרך החומצה האמינית האחרונה שהוספה. אתר A ריק כדי לקבל מולקולה נוספת.
להרחבה – Alberts, מהדורה שביעית, ע”מ 365-375
מולקולרית 4 שאלה 28
באאוקריוטים, כמה אנרגיה נחוצה לתרום מולקולת mRNA שהתוצר שלה הוא חלבון המכיל 4 חומצות אמינו (כולל החומצה הראשונה של הinitiator tRNA)?
תשובה ג‘. שאלה מעט מסובכת, אך היא באה להבהיר ולחדד את שימוש האנרגיה והפקטורים שפועלים בתהליך התרגום (Translation). פירוט השלבים:
אינציאציה (Initiation) – בתהליך ההטענה של ה-tRNA (מה שנקרא tRNA charging) משתמשים ב-ATP לכל tRNA אחד שמתחבר לח. אמינו מסוימת. מכיוון שה-ATP מפורק ל-AMP ופירופוספט הוא נחשב בסך האנרגיה כאילו פירקנו 2 ATP לשני ADP ולכן על כל ח. אמינו אחת מפורקים 2 ATP. סה”כ 8 ATP מכיוון שיש סה”כ 4 ח. אמינו בחלבון המדובר. בהמשך לזאת, יש שימוש ב-GTP אחד ההכנסה של ה-tRNA הטעון (Initiator tRNA) ע”י הפקטור eIF2 לתוך הריבוזום (לאתר P), ואחת על חיבור תתי-היחידות.
אלונגציה (Elongation) – על כל תזוזה של הריבוזום לקודון הבא (מה שנקרא Translocation) נעשה שימוש ב-GTP אחד ע”י הפקטור EF2. מכיוון שה-tRNA הראשון הוכנס ישירות לאתר P של הריבוזום ולא לאתר A (המקרה היחיד שבו זה קורה) אז בשביל להכניס את ה-tRNA השני של החלבון לא צריך לבצע טרנסלוקציה (כי אתר A פנוי מלכתחילה) ולכן נצטרך סה”כ 3 טרנסלוקציות של הריבוזום להכנסת 3 ח. האמינו הנוספות = כלומר 3 GTP שיפורקו ע”י EF2. בנוסף לזאת יש את הפקטור EF1 שמבצע פעילות דומה ל-eIF2 אשר מפרק GTP אחת על כל tRNA טעון שהוא מכניס לריבוזום ולכן הוא יפרק 3 GTP סה”כ בתרגום חלבון בעל 4 ח. אמינו (כי את הראשונה הכניס eIF2). סה”כ 3 טרנסלוקציות שזה 3 GTP ו-3 הידרוליזות של GTP ע”י EF2 שזה יחד 6 GTP.
טרמינציה (Termination) – לאחר הכנסת ח. האמינו הרביעית יכנס פקטור השחרור (Release factor) שהוא חלבון ולא tRNA אשר אינו משתמש בשום מטבע אנרגיה ויפיל את הריבוזום מה-mRNA (וזאת לאחר ביצוע הטרנסלוקציה הרביעית אשר תגרום לו להיכנס ל-P site = כלומר GTP אחד ע”י EF2). על מנת לפרק את תת-היחידה הגדולה מתת-היחידה הקטנה לאחר שחרור הריבוזום מה-mRNA (בשביל שיהיו מוכנים לעוד סיבוב של תרגום בהמשך) יגיע פקטור חלבוני אשר ישתמש ב-GTP אחד ויפריד ביניהם, ולכן נוסף פה עוד GTP אחד לסיום. סה”כ GTP אחד לטרנסלוקציה האחרונה שבה פקטור השחרור יעבור ל-P site ו-GTP נוסף כאשר יפורק הריבוזום ל-2 תת-היחידות (הקטנה והגדולה) = 2 GTP. שימו לב שהפירוט על הטרנסלוקציה האחרונה לא קיים באלברטס אלא רק הסקה מהאיור שבעמוד 349 בסילבוס.
לסיכום, באינציאציה יש שימוש ב-2 GTP ו-8 ATP. באלונגציה יש שימוש ב-6 GTP ובטרמינציה יש שימוש ב-2 GTP. סה”כ 8 ATP ו-10 GTP לתרגום של חלבון בעל 4 חומצות אמינו.
הערת המחבר – חשוב להכיר כי בתהליך הסריקה (Scanning או Searching) של תת-היחידה הקטנה בשלב האינציאציה יש שימוש בהליקאזות אשר מאפשרות את תנועתה, הליקאזות אלו משתמשות במספר של ATP עד להגעה לקודון ה-AUG הראשון. אנרגיה זו לא נספרה בחישוב סך האנרגיה. בנוסף לזאת, נאמר באלברטס כי באינציאציה יש עוד שימוש של GTP לפני חיבור תתי-היחידות של הריבוזום, אך לא מפורט ע”י מי ולמה, ולכן גם לא הוכנס לחישוב סך האנרגיה בתהליך התרגום.
להרחבה– Alberts, מהדורה שביעית, עמ’ 365-375 (Translation).
מולקולרית 4 שאלה 29
מה מהבאים אינו נכון בהקשר של תהליך התרגום של הריבוזום?
תשובה א‘. ישנם 2 מושגים חשובים שצריך להכיר בהקשר תהליך ההגהה (Proofreading) של הריבוזום. הראשון נקרא Induced fit והוא מתאר את הקיפול (ה”חביקה”)של הריבוזום על זיווג הבסיסים עוד לפני כניסת ח. האמינו לפוליפפטיד. כזכור ההבדלבין זיווג הבסיסים של T ו-A ל-G ו-C הוא בקשר מימן אחד (לראשונים 2 קשרי מימן ולאחרונים 3 קשרי מימן), ולכן כאשר לא תהיה התאמה נכונה בין הבסיסים תהייה הפרעה מרחבית שתפסיק את פעולתו של הריבוזום. בהמשך לזאת, המשוג השני שקשור לתהליך ההגה הנקרא Kinetic proofreading והוא מתאר את הזמן שלוקח לבצע הידרוליזה ל-GTP כזמן שבו הריבוזום מוודא שאכן הקודון תואם לאנטי-קודון, כלומר ישנוניצול של ההפסקה הקצרה בתהליך הקידוד לאיתור שגיאות. ה-Kinetic proofreading גורם להעלאת הספציפיות של זיווג הבסיסים מכיוון שהוא מבטל זיווגים לא ספציפיים (לא תואמים), ולכן מסיח זה אינו נכון.
שלילת מסיחים:
ב’. בפרוקריוטים, ה-16S שנמצא בתת-היחידה הקטנה אחראית על תהליך ה-Induced fit של הריבוזום והיא זו ש”חובקת” את זיווג הבסיסים, ולכן מסיח זה נכון.
ג’. כפי שנאמר מעל, ההגהה הקינטית קשורה בזמן שלוקח לבצע הידרוליזה ולכן מסיח זה נכון.
ד’. כמובן שה-Induced fit מתרחש לפני העברת ח. אמינו באתר A לפוליפפטיד באתר P מכיוון שזה בא לבדוק התאמה ונכונות של זיווג הבסיסים בשביל למנוע הכנסה שגויה של ח. אמינו ולכן מסיח זה גם נכון.
להרחבה – Alberts, מהדורה שביעית, עמ’ 365-368 (Ribosome) ועמ’ 369-371 (Induced fit + Kineticproofreading).
מולקולרית 4 שאלה 30
איזה מהמשפטים הבאים הוא הנכון?
תשובה ב‘. פקטור האיניציאציה האאוקריוטי eIF2 אחראי לקישור Aminoacyl-tRNA ההתחלתי (tRNAi) לריבוזום. פקטור זה דורש GTP לפעולתו שעובר הידרוליזה כשהפקטור מתנתק מקומפלקס האינציאציה. גם פקטורי האלונגציה (בפרוקריוטים ובאאוקריוטים) דורשים GTP כדי לבצע את עבודתם: EF-Tu מביא את מולקולת Aminoacyl-tRNA לאתר A בריבוזום, וכדי להשתחרר מהקומפלקס, מבצע הידרוליזה ל-GTP. EF-G מבצע הידרוליזה ל-GTP במהלך שלב הטרנסלוקציה (Translocation) של שתי תתי היחידות של הריבוזום (הגדולה והקטנה).
שלילת מסיחים:
א’. הקשר הפפטידי מתבצע על ידי פעילות הפפטידיל טרנספראז (Peptidyl transferase) שלהיחידה 23S בתת היחידה הגדולה בריבוזום הפרוקריוטי (וב-28S באאוקריוטים), ולא ע”י ה-16S שנמצאת בתת-היחידה הקטנה של הריבוזום הפרוקריוטי ולכן מסיח זה אינו נכון.
ג’. כפי שנאמר מעל, הטרנסלוקציה מתבצעת ע”י פקטור שנקרא EF-G (או EF-2 באאוקריוטים) ולכן מסיח זה אינו נכון. הפקטור EF-Tu אחראי להבאת מולקולת ה-tRNA המתאימה לריבוזום.
ד’. ה-tRNA היחיד שנכנס לאתר P הוא ה-tRNA ההתחלתי (tRNAi), לאחר מכן כל המולקולות הבאות יכנסו לאתר A בלבד. באתר P נמצא הפוליפפטיד שמסונתז.
להרחבה – Alberts, מהדורה שביעית, ע”מ 368-374 (Translation).
מולקולרית 4 שאלה 31
מה לא נכון לגבי RF (פקטור שחרור) בפרוקריוטים?
תשובה ד‘. פקטורי השחרור (Release factors) הם חלבונים אשר מסוגלים להתחבר לקודוני הסיום (Stop codons) ולהפסיק את התרגום (Translation). פקטורים אלו אינם סוג של tRNA (שמן הסתם בנויים מגדילי RNA) אלא בנויים מחלבונים, אך הם בנויים בצורה מרחבית מאוד דומה ל-tRNA מה שמקנה להם יכולת כניסה ל-A-site של הריבוזום ועקב כך לגרום להפסקת התרגום. כאשר פקטור השחרור נכנס לאתר A הוא גורם לאנזים פפטידיל טרנספראז (Peptidyl transferase) ל”חבר” מולקולת מים לפוליפפטיד במקום ח. אמינו מה שגורם לשיחרורו של הפוליפפטיד ולסיום התרגום של ה-mRNA. ולכן מסיח זה לא נכון ואותו נסמן.
שלילת מסיחים:
א’. ישנם שלושה קודוני סיום – UAA, UAG ו-UGA (אני אוהב לזכור אותם כ”או-אה”, “או-אג”ו-“עו-גה”) אשר אליהם מתחבר פקטור השחרור ולכן מסיח זה נכון.
ב’. כפי שנאמר מעל, ה-tRNA וה-Release factors חולקים מבנה מרחבי דומה ולכן מסיח זה גם נכון.
ג’. אכן פקטורי השחרור נכנסים לאתר A (כמו כל tRNA – מלבד ה-tRNAi הראשון).
להרחבה – Alberts, מהדורה שביעית, ע”מ 365+367 (Peptidyl transferase), ע”מ 374-375 (Release factor)