
שאלון מערכת: שיעורי בית ביוכימיה 4
ביוכימיה 4 שאלה 1
אדנילאט ציקלאז (adenylate cyclase) מופעל ע”י:
במה השאלה עוסקת: איתות תאי, איתות דרך Gs
תשובה ד. אדנילאט ציקלאז (Adenylate Cyclase) מופעל במסלול המתווך ע”י חלבוני G מסוג Gs. כאשר ליגנד נקשר לרצפטור בצד החוץ תאי, זה מוביל לשינוי בצד התוך תאי של הרצפטור, שם לרצפטור קשור טרימר של חלבון G (אלפא-בטא-גאמא). בעקבות הפעלת הרצפטור, תת יחידה אלפא, אשר קושרת GDP, עוברת שחלוף שבעקבותו תקשור GTP. דבר זה מוביל להתנתקות תת יחידה אלפא מתת יחידה בטא וגאמא. לאחר מכן, תת יחידה אלפא, שכרגע קושרת GTP, מפעילה את האנזים אדנילאט ציקלאז.
שלילת מסיחים:
א. אדנילאט ציקלאז אמנם מופעל ע”י תת יחידה αs אך תת יחידה זו קושרת GTP בעת הפעלת אדנילאט ציקלאז.
ב. האנזים PKC מזרחן מספר מטרות בתא במסלול הסיגנל של רצפטור מסוג Gq.
ג. רצפטור בטא אדרנרגי הוא דוגמא לרצפטור המתווך לחלבוני Gs. אדנילאט ציקלאז לא מגיב עם הרצפטור ישירות, אלא בתיווך של חלבון αs.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 896-898
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 2
כל המשפטים הבאים בנוגע לחלבוני G מונומרים נכונים, מלבד:
במה השאלה עוסקת: איתות תאי, חלבוני GTPase
תשובה ג. חלבוני G מונומרים משתתפים במגוון תהליכים בתא, כגון יצירת וזיקולות, אך אינם מבקרים אנזימים המייצרים cGMP. שליח שניוני זה מבוקר ע”י חלבוני G טרימרים או ע”י הגז NO. חלבוני G במצב הלא פעיל קושרים GDP, ועל מנת לעבור לצורתם הפעילה (קשירת GTP) הם זקוקים לסיוע של אנזים משחלף מסוג GEF (ר״ת: Guanine Nucleotide Exchange Factor). כחלק מהבקרה על פעילות חלבוני G, אנזים מסוג GAP (ר״ת: GTPase Activating Protein) מסייע בהידרוליזה של ה-GTP ל-GDP, וכך החלבון חוזר למצבו הלא הפעיל.
שלילת מסיחים:
א. כפי שרשום לעיל, חלבוני G מונומרים מבוקרים ע”י אנזימי GEF. אנזימים אלו מוציאים את מולקולת ה-GDP מחלבון ה-G ובמקום מכניסים לחלבון ה-G מולקולת GTP.
ב. כפי שרשום לעיל, חלבוני G מונומרים מבוקרים ע”י אנזימי GAP. לחלבוני G יש יכולת אינטריזית לפעילות GTPase אך היא חלשה ומתרחשת לאט יותר, ולכן אנזימי GAP מסייעים בכך.
ד. יצירת וזיקולות מתווכת ע”י חלבוני G מונומרים כדוגמת ARF.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 880
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מבוא לאיתות תאי
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 3
אדם הסובל מהיפוקסיה כרונית (רמות חמצן נמוכות), עקב ירידה בתפקוד הריאות יוכל להסתגל, ע”י העלאת רמות ה- BPG בדם. מה מהבאים נכון עבור אותו אדם?
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה ב. מולקולת 2,3BPG (ר״ת: 2,3-Bisphosphoglycerate) בעלת מטען שלילי, ולכן, בעת קישורה להמוגלובין היא יכולה ליצור קשרים אלקטרוסטטיים. נזכר כי קשרים אלקטרוסטטיים במולקולת ההמוגלובין מעודדים מצב T – קונפורמציה בעלת אפיניות נמוכה יותר לחמצן מאשר מצב R. ערך p50 הוא לחץ החמצן בו 50% מההמוגלובין קושר חמצן, וזהו גם המדד לזיקה בין ההמוגלבין לחמצן (Kd). ככל שערך זה נמוך יותר, כך זיקת ההמוגלובין לחמצן גבוהה יותר (כלומר, דרוש לחץ חמצן נמוך יותר בכדי להגיע למצב בו 50% מההמוגלובין קושר חמצן). לכן, בנוכחות 2,3BPG ערך p50 יעלה. כמו כן, במצב של היפוקסיה רמת החמצן ברקמות נמוכה, והעלאת ריכוז 2,3BPG יוביל לעלייה בשחרור החמצן ברקמות.
שלילת מסיחים:
א. ירידה בערך p50 משמעותה שיש עלייה בזיקה בין ההמוגלובין לחמצן. 2,3BPG מוריד את הזיקה של ההמוגלובין לחמצן ולכן מסיח זה נפסל.
ג. מולקולת 2,3BPG מעודדת מצב T, לו זיקה נמוכה לחמצן. מצב R הינו קונפורמציה בעלת זיקה גבוהה לחמצן.
ד. המוגלובין הינו חלבון בעל קופראטיביות חיובית, קישור של מולקולת חמצן לאחת מתת היחידות מעלה את הזיקה בשאר תת היחידות. קופראטיביות נותנת עקומת קשירה סיגמואידית. אפקטורים שליליים של המוגלובין מסיטים את העקומה ימינה (עליה בערך p50) אך אינם מבטלים את הקופראטיביות (ביטול הקופראטיביות מתרחש בהרעלת CO – מה שמוביל לעקומה היפרבולית).
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Oxygen Binding to Hemoglobin Is Regulated by 2,3- Bisphosphoglycerate
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 4
מי מבין ההיגדים הבאים נכון אודות אפקט בוהר והמוגלובין?
( I ) אפקט בוהר יסיט את עקומת רוויון החמצן לימין ככל שה- pH ירד.
( II ) אפקט בוהר יסיט את עקומת רוויון החמצן לימין ככל שה- pH יעלה.
( III ) אפקט בוהר גורם לשחרור חמצן ברקמות המבצעות נשימה תאית.
( IV ) חמצן ופרוטונים מתחרים על קישור המוגלובין.
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה א. אפקט בוהר מתייחס להשפעת הפד״ח וה- pH על קישור ושחרור החמצן מהמוגלובין. ברקמות פעילות מטבולית יש ייצור מוגבר של פרוטונים (המתקבלים בנשימה תאית) ומכאן שערך ה-pH יהיה נמוך יותר. פרוטונים אלו יכולים להיקשר לשיירים של חומצות אמיניות בחלבון, כגון היסטידין, מה שיוביל לשינוי המטען בהמוגלובין ויצירת קשרים אלקטרוסטטים. קשרים אלו מעודדים מצב T, מצב בעל זיקה נמוכה לחמצן, מה שיוביל לשחרור חמצן ברקמות. ככל שריכוז הפרוטונים גבוה יותר (pH נמוך יותר) כך הזיקה בין ההמוגלובין לחמצן תהיה נמוכה יותר, ועקומת הקשירה של המוגלובין תסטה ימינה.
שלילת מסיחים:
ב. משפט I נכון כפי שצוין, אך משפט IV שגוי. להמוגלובין יש אתר קישור מסוים לחמצן ואתר קישור אחר לפרוטונים, לכן החמצן והפרוטונים לא מתחרים על הקישור בהמוגלובין.
ג. משפט III נכון כפי שצוין, אך משפט II שגוי. עליה בסולם ה-pH משמעותה ירידה בריכוז הפרוטונים, ולכן תהיה זיקה גבוהה יותר בין ההמוגלובין לחמצן. במצב זה, העקומה תסטה שמאלה.
ד. משפט II ו-IV שגויים כפי שהוסבר.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Hemoglobin Also Transports H+ and CO2
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 5
מי מבין ההיגדים הבאים נכון באשר לרמות הרוויה (saturation) של קשירת סידן ע”י החלבון הרגולטורי קלמודולין ( calmodulin )?
במה השאלה עוסקת: קשירת חלבונים וליגנדים
תשובה ב. קבוע הדיסוציאציה הינו מדד לזיקה בין החלבון לליגנד. ככל שקבוע הדיסוציאציה יותר נמוך, כך דרוש ריכוז ליגנד נמוך יותר על מנת שחצי ממולקולות החלבון יקשרו את הליגנד. לכן, ככל שערך Kd קטן יותר כך הזיקה של קלמודולין לסידן גבוהה יותר.
שלילת מסיחים:
א. Y מתאר את פרקציית הקשירה (כמה חלבון קשר ליגנד), ולכן ככל שיותר מולקולות קלמודולין קשרו סידן כך תטא תהיה גבוהה יותר.
ג. קלמדולין הינו חלבון קופראטיבי המכיל ארבעה אתרי קישור לסידן. לכן, עקומת הקשירה של קלמודולין תהיה סיגמואידית ולא היפרבולית.
ד. כאשר Y שווה ל- 0.5, חצי מהקלמודולין קשורות לסידן, ולא להפך.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Protein-Ligand Interactions Can Be Described Quantitatively
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 5
ביוכימיה 4 שאלה 6
לפניכם שישה היגדים:
היגד 1 — חמצן הוא מודולטור אלוסטרי הומוטרופי חיובי של המוגלובין.
היגד 2 — קשירת החמצן להמוגלובין מצויה תחת שני סוגים עיקריים של בקרה: שיתופיות חיובית ובקרה אלוסטרית הומוטרופית.
היגד 3 — באנמיה חרמשית, תאי הדם האדומים הופכים לצורת חרמש עקב אגרגציה של מולקולות המוגלובין קושרות חמצן.
היגד 4 — חלבון עם אתר קשירה בודד לליגנד שלו, יוכל ״לייצר״ עקומת קישור סיגמואידית אך ורק אם קשירת הליגנד מעוררת שינוי קונפורמציה בחלבון.
היגד 5 — ללא נוכחות של BPG בתאי דם אדומים, עקומת קשירת החמצן של המוגלובין הופכת להיפרבולית.
היגד 6 — עבור חלבון מולטמרי המכיל מספר אתרי קישור לליגנד שלו – מקדם היל בהכרח יהיה גבוה מ- 1, ואילו לחלבון מונומרי עם אתר קישור יחיד לליגנד יהיה בהכרח nH=1.
סמנו את המסיח הנכון ביותר:
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה א. היגד 1 — נכון. שיתופיות חיובית היא צורה של קשירה אלוסטרית, ולכן חמצן יכול להיחשב גם כליגנד של Hb וגם כמודולטור הומטרופי חיובי.
היגד 2 — לא נכון. במקרה של Hb, שיתופיות חיובית ובקרה אלוסטרית הומוטרופית חיובית מתייחס לאותו דבר כפי שמוסבר לעיל. בתכלס יותר מדויק להגיד ששתי הסוגים העיקריים הם בקרה אלוסטורית הומוטרופית חיובית (שיתופיות חיובית) ובקרה אלוסטורית הטרוטרופית שלילית (כל העניין עם אפקט בוהר, BPG וכו׳).
היגד 3 — לא נכון. באנמיה חרמשית ההמוגלובין המוטנט נקרא HbS והמחלה נובעת ממוטציה בודדת: Val במקום Glu בעמדה 6 בתתי-יחידות בטא. ההחלפה הזו יוצרת נקודת מגע הידרופובית “דביקה” שהמוגלובין, ורק כאשר ה- HbS משחרר חמצן הוא הופך לבלתי-מסיס וגורם לאגרגציה: הנקודות הדביקות הללו מביאות להידבקות של מולקולות deoxyHbS בתוך התא דם אדום, וזה בעצם יוצר את האגרגטים הארוכים שגורמים לתאי דם אדומים להיות בצורת חרמש.
היגד 4 — לא נכון. חלבון מונומרי עם אתר קישור בודד לליגנד אינו יכול לייצר עקומת קישור סיגמואידית — גם אם הקישור מעורר שינוי קונפורמציה בחלבון — מכיוון שכל מולקולת ליגנד נקשרת באופן עצמאי ואינה יכולה להשפיע על קישור מולקולת ליגנד נוספת לחלבון כי אין עוד אתרי קישור להשפיע עליהם.
היגד 5 — לא נכון. כפי שראינו במצגת ביוכימיה 3 חלק א שקף 36, אמנם האפיניות הכללית של Hb לחמצן עולה משמעותית כאשר אין בכלל BPG בתא דם אדום והעקומה נראת הרבה פחות סיגמואידית, אך עדיין יש שיתופיות בקשירה (מקדם היל יהיה 2.5).
היגד 6 — לא נכון. החלק הראשון במשפט שגוי, כי nH (מקדם היל) שווה ל- 1 יכול לקרות גם במקרה של חלבון מולטמרי המכיל מספר אתרי קישור — אך אין תקשורת בין התתי-יחידות שלו. להזכירכם מקדם היל הוא מדד לקישור שיתופי (מעל 1 = שיתופיות חיובית, מתחת ל- 1 = שיתופיות שלילית, וערך שווה ל- 1 אין שיתופיות).
להרחבה — Lehninger מהדורה שמינית, פרק 5.1
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 7
מי מהמולקולות הבאות המשתתפות בהעברת אותות אינה משמשת כשליח שניוני (second messenger)?
במה השאלה עוסקת: איתות תאי, שליחים שניוניים
תשובה ב. הגדרת שליחים שניוניים (second messengers, נקראים גם small intracellular mediator) = מולקולת איתות תוך-תאית קטנה שנוצרת או משתחררת לפעולה בתגובה לסיגנל חוץ-תאי ועוזרת להעביר את הסיגנל בתוך התא.
• ה”שליח הראשוני” זה הסיגנל החוץ-תאי שנקשר לרצפטור.
• השליחים השניוניים שיש לזכור: cAMP , cGMP, Ca+2, IP3 ו- DAG ; חלקם מסיסים במים ומדפייזים בציטוזול, וחלקם מסיסים בשומן ומדפייזים במישור הממברנה הפלזמטית.
התגובה של תא האנדותל לסיגנל אצטילכולין – היא ייצור הסיגנל של תאי השריר החלק של כלי הדם – NO, מולקולה גזית קטנה והידרופובית אשר מדפייזת החוצה מתאי האנדותל ונכנסת לתאי השריר החלק הסמוכים של כלי הדם. שם, NO גורם להרפיית השריר החלק של כלי הדם ה- NO אינו נחשב שליח שניוני במסלול הזה, כי הוא לא מפיץ את הסיגנל בתוך התא שבו הוא מיוצר.
שלילת מסיחים:
א. סידן נחשב לשליח שניוני נפוץ בהעברת אותות. בתוך חלל ה-E.R יש מאגר של יוני סידן, ובתגובה לסיגנל הסידן משתחרר אל הציטופלזמה ומווסת חלבונים שונים.
ג. אינוזיטול 1,4,5 טריפוספסט (בשמו המקוצר – IP3) הינו שליח שניוני הנוצר מחיתוך של פוספוליפיד במסלול המתווך ע”י Gq.
ד. המולקולה cAMP נוצרת מאדנילאט ציקלאז (Adenylate Cyclase) בעת הפעלת רצפטור המצומד ל-Gs. המולקולה cAMP נחשבת לשליח שניוני.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 908-909
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 8
מה מהבאים נכון באשר להמוגולובין עוברי?
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה א. המוגלובין של אדם בוגר (HbA) מורכב משתי יחידות אלפא ושתי יחידות בטא. בעובר, ישנו המוגלובין הנקרא HbF (כלומר Fetal, המוגלובין עוברי) אשר מורכב משתי יחידות אלפא ושתי יחידות גאמא. הכיס הנוצר בין תת יחידה גאמא לאלפא (שזהו אתר הקישור של 2,3BPG) בעל אפיניות נמוכה יותר ל-2,3BPG (שזהו אפקטור אלוסטרי הטרוטרופי שלילי של המוגלובין) ולכן להמוגלובין עוברי יש זיקה גבוהה יותר לחמצן מאשר להמוגלובין בוגר.
שלילת מסיחים:
ב. להמוגלובין עוברי זיקה נמוכה יותר ל-2,3BPG ולכן זיקתו לחמצן גבוהה יותר.
ג. כאמור, המוגלובין עוברי קושר חמצן ביעילות גבוהה יותר מאשר המוגלובין בוגר. כמו כן, בלחצי חמצן מספיק גבוהים המוגלובין עוברי והמוגלובין בוגר יקשרו חמצן באותה יעילות.
ד. כאמור, המוגלובין עוברי זיקה גבוהה יותר לחמצן מאשר המוגלובין בוגר.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Oxygen Binding to Hemoglobin Is Regulated by 2,3- Bisphosphoglycerate
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 9
מי מבין המולקולות הבאות מושפעת באופן הישיר ביותר ע”י חנקן חד חמצני (NO)?
במה השאלה עוסקת: איתות תאי, NO
תשובה ג. חנקן חד-חמצני (NO) נוצר בתגובה לסיגנל בתא ומופרש מהתא בדיפוזיה. לאחר מכן, מגיע לתא שכן (נכנס בדיפוזיה פשוטה) ובתוך התא נקשר לאנזים גואנליאט ציקלאז (Guanylate Cyclase) אשר הופך מולקולת GTP ל-cGMP.
שלילת מסיחים:
א. הגז NO אינו משפיע על רמות ATP באופן ישיר.
ב. עליה ברמת הסידן בציטוזול מושפעת ממסלול שונים, כגון ע”י מסלול Gq.
ד. עליה ברמת IP3 בציטוזול מושפעת ממסלול Gq.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 908-909
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 10
מי מהבאים אינו מהווה חלק מתהליך כיבוי סיגנל ה- GPCR?
במה השאלה עוסקת: איתות תאי, GPCRs
תשובה ד. כחלק מכיבוי הסיגנל של רצפטור GPCR (בתהליך הדה-סנסיטיזציה), הרצפטור תחילה עובר זרחון ע”י קינאז (כדוגמת βARK במקרה של הרצפטור הבטא-אדרנרגי לאינסולין/אפינפרין). הזרחון מגייס את החלבון ארסטין, וארסטין מתווך את האינטרליזציה (אנדוציטוזה) של הרצפטור לתוך התא (ולא של תת יחידה אלפא השייכות לחלבון G). הרצפטור לאחר מכן יכול לחזור לממברנה או לעבור פירוק בליזוזום. תת יחידה אלפא איננה עוברת את הפירוק אלא הרצפטור עצמו.
שלילת מסיחים:
א. זרחון ע”י הקינאז βARK הינו השלב הראשון בכיבוי הסיגנל של GPCR כפי שהוסבר.
ג. גיוס ארסטין הינו השלב השני בכיבוי הסיגנל של GPCR כפי שהוסבר.
ד. אינטרליזציה של הרצפטור זהו השלב השלישי בכיבוי הסיגנל של GPCR כפי שהוסבר.
הרחבה:
כאשר תאי מטרה נחשפים לסיגנל חוץ-תאי למשך תקופה ממושכת, הם יכולים לאבד רגישות לסיגנל. סוג חשוב של מנגנוני אדפטציה תלוי ב״טיפול ברצפטור עצמו״ — כלומר חסימת הרצפטור מפעולה או הורדת מספר הרצפטורים.
דה-סנסיטיזציה של GPCRs תלויה בזרחון הרצפטור ע״י קינאזות כמו PKA ו- PKC, או ע״י חבר במשפחת ה- GRKs (ר״ת: GPCR kinases). ה- GRKs מזרחנים מספר ח״א סרין ותראונין ב- GPCR, אך הם עושים זאת רק לאחר שקישור של ליגנד הפעיל את הרצפטור (כי הרצפטור המופעל עצמו הוא זה שמאקטב את ה- GRK באופן אלוסטרי). כמובן שגם זרחון ע״י PKA/PKC קורה רק כשהרצפטור פעיל.
ברגע שרצפטור עבר זרחון, הוא נקשר בזיקה גבוהה לחבר במשפחת חלבוני הארסטין — וכעת יש 3 אופציות:
(1) השבתת הרצפטור = הארסטין הקשור ל- GPCR מונע ממנו ליצור אינטראקציה עם החלבוני G.
(2) בידוד הרצפטור = הארסטין יכול לתפקד כאדפטור הגורם לאנדוציטוזה תלויות-קלתרין, כך שהרצפטור נכנס לתוך התא (אינטרליזציה) באופן זמני (בדיוק כמו שראינו בשקף הקודם עם הרצפטור הבטא-אדרנרגי).
(3) הרס הרצפטור = אותו דבר כמו (2) רק שזה לא יהיה זמני והרצפטור לא יחזור לממברנה, אלא יפורק בליזוזומים לאחר האינטרליזציה (מה שנקרא דאון-רגולציה).
הערה: אנדוציטוזה של הרצפטור אינה בהכרח עוצרת את האיתות. במקרים מסוימים, הארסטין שקשור לרצפטור מגייס חלבוני איתות אחרים כדי להעביר את הסיגנל הלאה מה- GPCRs המופנמים (internalized) לאורך מסלולי איתות חדשים .
להרחבה – Alberts, מהדורה שביעית, עמ׳ 909-910
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 11
מי מהבאים מפעיל מסלול איתות שכלל לא תלוי ב-cAMP ?
במה השאלה עוסקת: איתות תאי
תשובה א. הרצפטור לאינסולין הינו טירוזין קינאז. בעת היקשרות אינסולין לרצפטור, הצד התוך תאי של הרצפטור עובר אוטופוספורילציה על גבי שיירי טירוזין. לשיירי הטירוזין נקשר החלבון Grb2 המשמש כחלבון מתווך לחלבון SOS (ה-GEF של Ras) ואליו נקשר Ras אשר מפעיל את מסלול MAPK למשל.
שלילת מסיחים:
ב. הרצפטור לאדרנלין הינו רצפטור המתווך לחלבון Gs. מסלול Gs מפעיל אדנילאט ציקלאז אשר מייצר cAMP.
ג. אודורנטים הם מולקולות ריח, והרבה מהם מפעילים רצפטור המתווך לחלבון Golf, לו תת יחידה אלפא אשר מפעילה אדנילאט ציקלאז שמייצר cAMP.
ד. הורמון זה מפעיל מסלול איתות התלוי בעלייה ב- cAMP (כפי שניתן לראות בטבלה).

להרחבה – Alberts, מהדורה שביעית, עמ׳ 895-896
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 12
איזה רכיב איתות יכול להיות מופעל על-ידי RTKs אך לא על-ידי GPCRs?
במה השאלה עוסקת: איתות תאי
תשובה ד. Grb2 מתפקד כחלבון מתאם בחלק ממסלולי האיתות של RTKs. חלבון זה מכיל דומיין SH2 שדרכו הוא נקשר לפוספוטירוזין הקיים על הרצפטור עצמו, או במקרה של הרצפטור לאינסולין — על החלבון IRS1 (שנקשר לפוספוטירוזין על הרצפטור ואז מזורחן ע״י הרצפטור על טירוזין). לכן זה הגיוני ש- GPCR לא יוכלו להפעיל את Grb2 כי אין להם פוספוטירוזין.
שלילת מסיחים:
מסיח א — PLC מבקע את PIP2 בממברנת התא לשני שליחים שניוניים: DAG ו- IP3 (שגורם לעליית ריכוז הסידן הציטוזולי ע״י פתיחת תעלות ב- ER). ישנם כמה סוגים של PLC (כולם עושים די אותו דבר): GPCRs מפעילים את PLC-בטא ו- RTKs מפעילים את PLC-גמא.
מסיח ב — קיימים סוגים שונים של PI 3-kinases, אלה שמאוקטבים ע״י RTKs ו- GPCRs שייכים למחלקה I: קולטני RTKs משפעלים PI3Ks ממחלקת Ia, ו- GPCRs משפעלים PI3Ks ממחלקת Ib.
מסיח ג — GPCRs מסוימים יכולים גם הם להפעיל את Ras (ובכך את קסקדת ה- MAPK), אך הם עושים זאת ללא תלות באדפטור Grb2, אלא באמצעות Ras-GEF שמופעל ע״י השליחים השניוניים +Ca2 ודיאצילגליצרול (DAG).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 920-923
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 13
מי מהבאים שגוי?
במה השאלה עוסקת: איתות תאי, בקרת משוב
תשובה ב. אם מתרחש משוב חיובי במסלול איתות בעוצמה חזקה מספיק, המשוב יכול לייצר תגובת all-or-none. תגובת הכל-או-כלום כזו הולכת יד ביד עם תכונה נוספת: ברגע שהמערכת המגיבה עברה לרמת ההפעלה הגבוהה – המצב הזה בדר״כ משמר ומקיים את עצמו, משמע הוא יכול להימשך גם לאחר שעוצמת הסיגנל יורדת חזרה מתחת לערכו הקריטי. במקרה כזה, המערכת אמורה להיות bistable: היא יכולה להתקיים במצב “כבוי” או במצב “מופעל”, וגירוי חולף (שיש לו משוב חיובי חזק מספיק) יכול להפוך אותה ממצב אחד לשני.
בניגוד למשוב חיובי, משוב שלילי מנטרל את השפעת הגירוי ובכך מקצר ומגביל את רמת התגובה, מה שהופך את המערכת לפחות רגישה להפרעות. לעיתים לולאת משוב שלילי מכילה מנגנון דיליי כלשהו שמאט את אות המשוב דרך הלולאה, כלומר לוקח קצת זמן לכל ״סבב״ של המשוב השלילי להתבטא.
משוב שלילי כזה עם דיליי ארוך יכול לייצר תגובות עם תנודות {אוסילציות}.
אם משוב שלילי פועל בדיליי קצר, המערכת מייצרת תגובה קצרה לסיגנל, אך התגובה דועכת במהירות גם בזמן שהגירוי נמשך. אם הגירוי מוגבר עוד יותר – המערכת מגיבה שוב בעוצמה, אבל עדיין התגובה דועכת במהרה. זוהי בעצם תופעת האדפטציה/ דה-סנסזיטיזציה, כלומר המנגנון המאפשר לתא להתאים את הרגישות שלו לסיגנל וכך תתקבל תגובה חולפת ובעוצמה נורמלית גם בנוכחות של גירוי חזק וממושך.
מנגנון דה-סנסיטיזציה מחייב שרכיב כלשהו של מערכת האיתות ייצור סיגנל מעכב מושהה שמפחית את עוצמת התגובה (=משוב שלילי בדיליי ארוך).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 888-891
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מבוא לאיתות תאי
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 14
איזה מהמאפיינים הבאים מתאים לרצפטורים לאינסולין?
תשובה ב. הרצפטור לאינסולין הינו רצפטור טירוזין קינאז. כאשר רצפטור זה מופעל הוא עובר אוטופוספורילציה, זרחון עצמי על גבי שיירי טריוזין בצד התוך תאי של הרצפטור. שיירי הטירוזין המזורחנים מהווים אתרי קישור לחלבוני סיגנל שונים.
שלילת מסיחים:
א. פעילות ATPase הינה פירוק ATP ל-ADP. פעילות זו אינה מתרחשת ברצפטור טירוזין קינאז.
ג. שבעה דומיינים טרנסממברנלים זהו מאפיין של קולטני GPCRs, והרצפטור לאניסולין הוא מסוג טירוזין קינאז (RTK).
ד. רוב רצפטורי טירוזין קינאז עוברים דימריזציה בעקבות קישור הליגנד לכל תת יחידה של הרצפטור. הרצפטור לאינסולין, בניגוד לרוב רצפטורי טירוזין קינאז נמצא כבר בצורה האינטגרלית שלו. עם זאת, הרצפטור לאינסולין מופעל רק כאשר הליגנד נקשר לרצפטור.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 911-912
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 15
מהו אופן הפעולה של חנקן חמצני (nitric oxide)?
במה השאלה עוסקת: איתות תאי
תשובה ב. הגז NO הינו מולקולה קטנה ודי הידרופובית. תכונה זו מאפשרת ל-NO לחצות ממברנה בדיפוזיה פשוטה, ולהכנס לתאים סמוכים בדיפוזיה סמוכה גם כן. מולקולה המופרשת מתא אחד ומשפיעה על תא שכן נחשבת כפועלת במסלול פאראקריני.
שלילת מסיחים:
א. אנדוקריני זו הפרשת הורמון לזרם הדם, ומזרם הדם ההורמון יגיע אל אתר המטרה שלו. צורה זו אינה מאפיינת את הגז NO.
ג. ג’וקסטאקריני זו הפעלה תלוית מגע. הפעלה זו מתרחשת כאשר לממברנת תא אחד קשור הליגנד, ועל גבי ממברנת תא אחר קשור הרצפטור. כאשר תאים אלו יבואו במגע הליגנד ייקשר לרצפטור.
ד. אקסוקריני זו הפרשה אל מחוץ לגוף, כגון הפרשת זיעה והפרשת אנזימי הלבלב לצינור המעי. הגז NO משפיע על תאים שכנים ברקמות בו נמצא.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 908
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 16
איזה רצפטור מפעיל מסלול איתות שאינו תלוי ב- Ras superfamily?
במה השאלה עוסקת: איתות תאי
תשובה א. ה- Ras superfamily מורכבת ממספר משפחות של GTPases מונומריים, אך רק החברים במשפחת Ras ובמשפחת Rho עוזרים להעביר סיגנלים מרצפטורי איתות בפני שטח התא.
רודופסין הוא GPCR המצוי במערכת הראייה. למרות שהספר מזכיר ש- GPCR מסוימים יכולים להפעיל את Ras, רודופסין לא מפעיל איזשהו חלבון GTPase מונומרי אלא הוא גורם להפעלת cGMP-פוספודיאסטראז (דרך Gt).
באופן כללי, RTKs רבים פועלים בשתי מנגנוני איתות תוך-תאיים עיקריים:
הם גורמים להפעלת GEFs שמפעילים GTPases מונומריים (Ras ו- Rho) כדי להעביר את הסיגנל בתוך התא.
הם גורמים להפעלת PI3K שמזרחן פוספואינוזיטידים בממברנת התא ליצירת PIPs שונים, כאשר החשוב שביניהם זה PI(3,4,5)P3 שדרכו בסופו של דבר מופעל הסרין/תראונין קינאז Akt (שנקרא PKB בלנינג׳ר). למשל הרצפטור לאינסולין מפעיל את IRS1 וכך מתפצלים ממנו שני הסוגים של מסלולי האיתות הללו (בכללי שני סוגי המסלולים הללו נפוצים בהרבה RTKs של פקטורי גדילה).
שלילת מסיחים:
מסיח ב — כאשר המיטוגן (ופקטור הגדילה) EGF נקשר ל- RTK שלו, הוא מפעיל את Ras שבתורו מפעיל את קסקדת ה- MAPK ובכך מניע את התא להתחלק.
מסיח ג — במצגת יש דוגמה כיצד RTKs ממשפחת ה- Eph (כלומר רצפטורים לאפרינים) גורמים להפעלה של Rho. בדוגמה ה- RTK מצוי בחרוט הגדילה בקצה אקסון נודד של נוירון מוטורי, וקישור הרצפטור לאפרין גורם להפעלת מסלול איתות שבסופו של דבר יביא לקריסת חרוט הגדילה של האקסון כדי לנווט אותו לעבר שריר המטרה הנכון שהנוירון אמור לעצבב.
מסיח ד — אינטגרינים הם סוג אחר של רצפטורים (הרחבה בביותא 4) הפועלים ע״י קשירת טירוזין קינאז ציטופלזמי שנקרא FAK. כאשר האינטגרין קושר ליגנד (איזשהו חלבון במטריקס החוץ-תאי), מולקולות של האנזים FAK נקשרות לזנבות הציטוזוליים של האינטגרינים המקובצים ומזרחנים זה את זה. הפוספוטירוזינים מהווים אתר עגינה לטירוזין קינאז ציטופלזמי נוסף שנקרא Src. לאחר מכן Src ו- FAK מזרחנים זה את זה ומזרחנים חלבונים אחרים שמתאספים בצומת, כולל רבים מחלבוני האיתות המשמשים RTKs, ואכן אינטגרינים יכולים לגרום להפעלה של Ras בדרך זו.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 905-906 (רודופסין), 915-920 (קולטני RTK) ו- 923-924 (אינטגרינים).
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 17
תא מטופל בכמות קטנה של ליגנד חוץ-תאי המעורר GPCR המצומד ל- Gq. מה הכי סביר שיקרה?
במה השאלה עוסקת: איתות תאי, סידן כשליח שניוני
תשובה ג. זו שאלה לא פשוטה, ראשית ננסה לעשות סדר — Gq גורם משפעל את PLC שבתורו מבקע את PIP2 ל- DAG ו- IP3. בממברנת ה- ER יש שני סוגים של תעלות סידן:
1. רצפטור IP3 — דורש קישור גם של IP3 וגם של סידן כדי שהתעלה תפתח.
2. רצפטור ריאנודין — נפתח בתגובה לעלייה מתונה בריכוז הסידן בציטוזול.
שני הרצפטורים הללו עוברים משוב חיובי בריכוזי סידן ציטוזוליים נמוכים עד בינוניים, ושניהם עוברים משוב שלילי (סגירת תעלה) בריכוזי סידן גבוהים.
אם התא מטופל בכמות קטנה של ליגנד, תהיה עלייה קטנה בריכוז ה- IP3 הציטוזולי שתוביל להתפרצות זעירה בריכוז הסידן הציטוזולי עקב פתיחת מספר קטן של רצפטורי IP3 בממברנת ה- ER שקשרו גם IP3 וגם +Ca2, כאשר הריכוזים של שניהם נמוכים מידי כדי להיקשר לכל הרצפטורים הללו (שלילת מסיחים א וד).
לעומת זאת, אם הסיגנל החוץ-תאי חזק יותר, IP3 עולה לריכוז גבוה יותר ונקשר להמון רצפטורי IP3 ב- ER. כעת, הפרץ המקומי הקטן של שחרור +Ca2 יכול להתפשט לרצפטורים שכנים שגם קושרים IP3 ולהפעיל אותם, וכתוצאה מכך נוצר גל רגנרטיבי של שחרור +Ca2 בציטוזול. רק במקרה כזה נראה תנודתיות בריכוז הסידן עקב לולאות המשוב שעוברים רצפטורי IP3 וריאנודין: עלייה קטנה בריכוז הסידן הציטוזולי מובילה לעלייה גדולה בריכוז הסידן (משוב חיובי) ← אחר כך מגיע משוב שלילי (בדיליי) עקב ריכוז סידן ציטוזולי גבוה ← הסרת המשוב השלילי ע״י משאבות סידן ← המעגל חוזר חלילה.
הרחבה על CaM-kinase II:
כאשר ריכוז הסידן הציטוזולי עולה, הסידן נקשר לקלמודולין ויחד הם מבקרים את הפעילות של חלבונים רבים בתא. אחד מהם הוא CaM-kinase II, המכיל 12 תתי-יחידות שלכל אחת פעילות של קינאז. האנזים הזה תלוי בסידן באופן הבא: דומיין הקינאז של כל תת-יח׳ מעוכב וכאשר +Ca2/קלמודולין נוכח בתא, הוא יכול להיקשר לתת-יח׳ ולמנוע את עיכוב דומיין הקינאז. אם זה קורה גם בתת-יח׳ הסמוכה, שני דומייני הקינאז יזרחנו זה.
ברגע שתת-יחידת קינאז עוברת אוטופוספורילציה בדרך זו, היא נשארת פעילה גם בהיעדר +Ca2 (כלומר לאחר שהקלמודולין-סידן התנתקו מהתת-יח׳), ובכך מאריכה את משך פעילות הקינאז מעבר למשך הזמן של סיגנל הסידן ההתחלתי ששיפעל אותו. האנזים שומר על פעילות זו עד שפוספטאז מסיר את האוטופוספורילציה ומכבה את הקינאז.
שלילת מסיח ב — כפי שהוסבר בהתחלה, רק כאשר יש עלייה גדולה בריכוז ה- IP3 נראה תנודות גדולות בריכוז הסידן הציטוזולי, ורק כאשר תדירות התנודות הללו היא גבוה יחסית, האנזים יוכל להגיע למצב שכל 12 תתי היחידות שלו פעילות ועברו אוטופוספרילציה.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 899-904
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 18
מי מההבאים אינו סוג של רצפטור הקשור לטירוזין-קינאז?
במה השאלה עוסקת: איתות תאי
תשובה ד. כידוע, Enzyme-coupled Receptors זה סוג כללי של רצפטורים המכילים פעילות אנזימתית אינטרנזית (כמו פעילות הטירוזין-קינאז של RTKs) או מצומדים לאיזשהו אנזים שעושה בשבילם את העבודה.
סוג אחד של רצפטורים כאלה אלו רצפטורים שתלויים בזרחון טירוזין לצורך פעילותם ובכל זאת חסרים דומיין טירוזין-קינאז אינטרינזי. רצפטורים אלה פועלים באמצעות אנזימי טירוזין-קינאז ציטופלזמיים (כמו למשל Src, FAK, JAK ועוד) הקשורים לרצפטורים ומזרחנים חלבוני מטרה שונים, לרוב כולל הרצפטורים עצמם, כאשר הרצפטורים קושרים את הליגנד שלהם.
חלבוני סיגנל חוץ-תאיים ממשפחת העל TGF-בטא הם הליגנדים של קולטני סרין/תראונין-קינאז (כמו קולטני RTK רק שקולטנים אלו מזרחנים סרין ותראונין ולא טירוזין).
שלילת מסיחים:
מסיחים א +ג — המחלקה הגדולה והמגוונת ביותר של רצפטורים המסתמכים על אנזימי טירוזין-קינאזות ציטופלזמיים היא קולטני ציטוקינים, וקולטנים אלו מפעילים את מסלול האיתות JAK–STAT. הטירוזין קינאז הציטופלזמי הקשור לרצפטור נקרא JAK, וחלבוני STAT הם בקרי שעתוק רדומים בציטוזול שנכנסים לגרעין לאחר שהם מופעלים באמצעות הזרחונים שמבצע JAK (והוא עצמו מופעל רק כאשר הקולטן קושר ציטוקין כמובן).
מסיח ב — אינטגרינים גם פועלים דרך קשירה של טירוזין-קינאז ציטופלזמי (הקינאז FAK) אשר נקשר לזנב הציטוזולי של האינטגרינים כאשר הם קושרים ליגנד.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 923-927
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – סוגים אחרים של רצפטורים המצומדים לאנזים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 19
מי מהבאים שגוי לגבי הפרוטאין-קינאז mTOR (ר״ת: mammalian target of rapamycin)?
במה השאלה עוסקת: איתות תאי, mTOR
תשובה ד’. בתאי יונקים, הקינאז mTOR קיים בשני קומפלקסים מרובי-חלבונים שונים:
הראשון הוא mTORC1 (ר״ת: mTOR complex 1), המכיל את החלבון raptor. קומפלקס זה רגיש לרפמיצין, והוא ממריץ את גדילת התא. שני האקטיבטורים המוכרים ביותר שלו הם פקטורי גדילה (שמפעילים את מסלול PI3K-Akt) ונוטריינטים כמו חומצות אמינו, שניהם בעצם מאקטבים את mTORC1 ובכך מקדמים גדילת תאים.
השני הוא mTORC2 (ר״ת: mTOR complex 2), המכיל את החלבון rictor ואינו רגיש לרפמיצין. mTORC2 עוזר לקדם את הישרדות התא ע״י סיוע בשפעול של Akt עצמו (במסלול PI3K-Akt) והוא גם מווסת את שלד האקטין באמצעות חלבוני GTPases מונומריים ממשפחת Rho.
מסיח ד שגוי מכיוון שהמסלול מקדם את הישרדות התא ע״י ביטול העיכוב של החלבון האנטי-אפופטוטי Bcl2. כדי שזה יקרה יש להפעיל את Akt, ו- mTORC2 תורם להפעלה שלו ובכך תורם להישרדות התא. שימו לב שמסלול PI3K-Akt יכול לקדם גם את גדילת התא ע״י סיוע בשפעול של mTORC1.
באיור מוצג כיצד מסלול PI3K-Akt מקדם את הישרדות התא:
סיגנל הישרדות חוץ-תאי נקשר ומפעיל RTK, אשר מגייס ומשפעל את PI3K.
כעת PI3K מייצר PI(3,4,5)P3 (ע״י זרחון של PI(4,5)P2) המשמש כאתר עגינה לשני קינאזות המכילים דומיין PH – הקינאז Akt והקינאז PDK1 – ובכך מביא אותם לקרבה זה לזה בממברנה הפלזמטית.
הקינאז Akt מזורחן ע״י mTORC2, מה שמשנה את הקונפורמציה של Akt כך שכעת הוא יכול לעבור זירחון נוסף ע״י PDK1, ופעולה זו מאקטבת את Akt.
הקינאז Akt הפעיל מתנתק מהממברנה ומזרחן מגוון חלבוני מטרה, ביניהם החלבון הפרו-אפופטוטי Bad (ממשפחת BH3-only). כאשר Bad אינו מזורחן, הוא מחזיק את החלבון אנטי-אפופטוטי Bcl2 במצב לא- פעיל.
לאחר הזרחון, Bad משחרר את Bcl2, אשר כעת יכול לחסום אפופטוזיס ובכך לקדם את הישרדות התא.
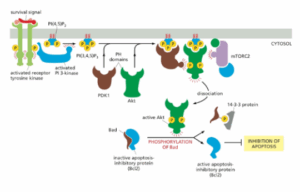
להרחבה – Alberts, מהדורה שביעית, עמ׳ 921-922
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 20
התאם בין ההיגד למושג המתאים לו בצורה הטובה ביותר: תתי יחידות אלפא וגמא, של חלבוני G הטרוטרימרים, מעוגנות לממברנת התא ע”י קישור קוולנטי לסוג זה של מולקולות.
במה השאלה עוסקת: איתות תאי, GPCR
תשובה ב’. תתי-יחידות אלפא וגמא של חלבוני ה- G הטרימריים מקושרות קוולנטית לליפידים ובכך מעוגנות לממברנה:
הקצה האמיני של אלפא קשור לפלמיטואיל, ובחלק מהמקרים גם למיריסטואיל.
הקצה הקרבוקסלי של גמא (כמו גם ה- GTPase המונומרי Ras) מקושר לאיזופרניל.
שלילת מסיחים:
א. קלמודולין מופעל במסלולי סיגנל בעקבות היקשרות של יוני סידן אליו. הוא אינו מעגן חלבוני G לממברנת התא.
ג. האנזים PKC מופעל בהעברת סיגנל של GPCR מסוג Gq ואינו מעגן את חלבוני ה-G לממברנה.
ד. אדרנלין (אפינפרין) הוא שליח ראשוני אשר נקשר לרצפטור GPCR ומפעיל אותו. הוא אינו מעגן את חלבוני G לממברנת התא.
להרחבה — Lehninger מהדורה שמינית, פרק 12.1 — G Proteins Act as Self-Limiting Switches in Many Processes
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 21
התאם בין ההיגד למושג המתאים לו בצורה הטובה ביותר: אנזים זה הופך לפעיל כאשר קשור ליוני סידן ודיאצילגליצרול (DAG).
במה השאלה עוסקת: איתות תאי
תשובה ג’. האנזים PKC עובר הפעלה ע”י קישור לדיאצילגליצרול (DAG) ויוני סידן. כאשר מופעל רצפטור GPCR מסוג Gq מופעל PLC המבקע פוספוליפיד (PIP2) בממברנה ל-DAG ו-IP3. השליח IP3 מוביל לשחרור יוני סידן מה-ER אל הציטוזול, והעלייה הראשונית בריכוז הסידן משנה את ה- PKC כך שהוא עובר מהציטוזול לפני השטח הציטופלזמי של הממברנה הפלזמטית. שם PKC משופעל על-ידי שילוב של +Ca2, דיאצילגליצרול והפוספוליפיד הממברנלי הטעון שלילית פוספטידיל-סרין.
שלילת מסיחים:
א. קלמודולין מופעל ע”י הקשרות של יוני סידן ואינו מופעל ע”י DAG. בנוסף, קלמודולין הוא אינו אנזים אלא מפעיל חלבונים באמצעות מגע.
ב. גואנליל-ציקלאז נמצא בשתי גרסאות: אנזים טרנסממברנלי המתפקד כרצפטור, והוא מופעל כאשר נקשר ליגנד לדומיין החוץ-תאי שלו. הגרסה השנייה זה אנזים מסיס בציטוזול, שהכרנו אותו במסלול ה- NO: אצטילכולין משפעל GPCR המצומד ל- Gq בתאי אנדותל ← העלייה בסידן גורמת לשפעול NOS ← ה- NO שמסונתז מדפייז לתוך תאי השריר החלק של כלי הדם ונקשר לגוואנליל-ציקלאז ← האנזים משופעל ומייצר cGMP (מה שמוביל להרחבת כלי הדם).
ד. השפעול הראשוני של CaM-kinase II קורה כאשר קלמודולין (במצבו הקושר סידן) נקשר אליו.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 898-899
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 22
לפניכם ארבעה היגדים על משפחת הגלובינים:
היגד 1 — גשרי מלח ממלאים את התפקיד העיקרי בייצוב הממשקים אלפא-בטא בתתי היחידות של המוגלובין.
היגד 2 — בזכות ההיסטדין הפרוקסימלית של מיוגלובין (His F8), יש הגדלה סלקטיבית של האפיניות לחמצן על פני CO.
היגד 3 — על אף שחמצן מולקולרי (O2) יכול לעבור בכוחות עצמו בזרם הדם, הוא מובל ע״י נשא המוגלובין כדי שהיה שחרור מבוקר שלו לרקמות.
היגד 4 — בידוד של מיוגלובין במבחנה עם תנאים שמורידים את ה- Kd (או p50) שלו לחמצן, משמעותו שבלחץ חלקי נמוך יותר של חמצן נקבל Y=0.5.
סמנו את התשובה הנכונה ביותר:
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין ומיוגלובין
תשובה א׳. היגד 1 — לא נכון (שלילת מסיח ב). מדובר בהיגד רשע ומבלבל: אמנם באזור הממשק ישנם גם גשרי מלח וקשרי מימן, אך אינטראקציות הידרופוביות ממלאות את התפקיד המרכזי בייצוב אזור המגע (הממשק) בין תתי-יח׳ אלפא-בטא של טטרמר ההמוגלובין, וכמובן שהוא נשאר טטרמר יציב גם במצב T וגם במצב R. התפקיד החשוב של גשרי מלח זה יותר בייצוב מצב T של המוגלובין ובמעבר בין מצב T ל- R המערב מספר שינויים בגשרי המלח.
היגד 2 — לא נכון. זה קורה בזכות ההיסטדין הדיסטלית (His E7), שמבצעת קשר מימן עם החמצן הקשור לאטום הברזל במרכז קבוצת ה- Heme.
היגד 3 — לא נכון (שלילת מסיח ד). חמצן לא יכול לעבור בכוחות עצמו בדם מכיוון שאינו מסיס במים.
היגד 4 — נכון (שלילת מסיח ג). הערך של Kd מייצג את ריכוז הליגנד שבו נקבל Y=0.5, כלומר הלחץ החלקי של חמצן שבו חצי מהמיוגלובין יקשור חמצן. לכן זה הגיוני שירידה ב- Kd משמעותה אפיניות גבוהה יותר, כך שאפשר לספק פחות חמצן במבחנה כדי שמחצית ממולקולות המיוגלובין יקשרו חמצן.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 23
הפעלה של Gq מובילה לכך שקלמדולין יעבור בין מספר קונפרמציות היפותטיות (המתוארות באמצעות צורות) בסדר הבא:
עיגול ← משולש ← ריבוע ← עיגול.
מה הכי סביר שכל קונפורמציה מייצגת?
במה השאלה עוסקת: איתות תאי, סידן כשליח שניוני
תשובה ב׳. חלבונים קושרי–+ Ca2 רבים מסייעים להעביר את סיגנל הסידן הציטוזולי. החשוב ביותר מבינהם הוא קלמודולין, המכיל ארבעה אתרי קישור לסידן. כאשר קלמודולין קושר סידן (שני יוני סידן לפחות), הוא עובר שינוי קונפורמציה משפעל. ההפעלה האלוסטרית של קלמודולין ע״י + Ca2 מקבילה להפעלה של PKA ע״י cAMP, אלא שלקומפלקס +Ca2/קלמודולין המאוקטב אין פעילות אנזימטית משל עצמו, והוא בעצם פועל באמצעות קישור והפעלה של חלבונים אחרים. כאשר +Ca2/קלמודולין נקשר לחלבון המטרה שלו – קלמודולין שוב משנה את הקונפורמציה שלו, ואופי השינוי תלוי בחלבון המטרה הספציפי שאליו הוא נקשר.
לפיכך, אם הסדר הנתון הוא עיגול ← משולש ← ריבוע ← עיגול (ראו איור מאולתר):
עיגול = הקונפרמציה של קלמודולין במצבו הנטול-סידן.
זמן קצר לאחר ש- Gq מופעל, ריכוז הסידן הציטוזולי עולה ולכן משולש = הקונפרמציה של קלמודולין לאחר שהוא קושר סידן (+Ca2/קלמודולין).
לאחר מכן קלמודולין נקשר לחלבון מטרה כלשהו שהוא מבקר (כמו למשל CaM-קינאז) ומשנה שוב את הקונפרמציה, ולכן ריבוע = +Ca2/קלמודולין הקשור לחלבון מטרה.
כאשר ריכוז הסידן הציטוזולי יורד, סידן יתנתק מקלמודולין כך שקלמדולין יתנתק מחלבון המטרה ויחזור לקונפרמצייה המקורית שלו (עיגול).
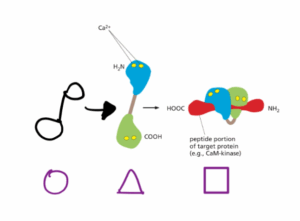
להרחבה – Alberts, מהדורה שביעית, עמ׳ 902
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 24
סמנו את המשפט השגוי
במה השאלה עוסקת: איתות תאי, RTKs
תשובה א’. אמנם שני סוגי הרצפטורים עוברים אנדוציטוזה, אך האנדוציטוזה מפסיקה את האיתות של הרצפטור ל- EGF ואילו במקרה של TrkA (=הרצפטור ל- NGF) זה מייעל את האיתות שלו.
הסבר:
כידוע, כאשר RTKs פעילים, לרוב נבנה על הזנב הציטוזולי שלהם קומפלקס איתות (שמתחיל מחלבונים המכילים דומיין ה- SH2 שנקשרים לפוספוטירוזינים על הרצפטור). עם זאת, לא כל החלבונים עוזרים להעביר את הסיגנל הלאה וחלקם פועלים כדי להפחית את תהליך האיתות, לדוגמה:
החלבון c-Cbl נקשר לכמה RTK פעילים (כמו הרצפטור ל- EGF) ומזרז את היוביקוויטילציה שלהם. חלבונים אנדוציטים המכילים UIMs מזהים את היוביקוויטילציה של הרצפטור ומכוונים אותו לוזיקולות מצופות-קלתרין, ובסופו של דבר פירוק של הרצפטורים בליזוזומים (= דאון-רגולציה).
אנדוציטוזה המושרה על-ידי ליגנד של RTKs לא תמיד מפחיתה את האיתות. במקרים מסוימים, RTK מופעלים עוברים אנדוציטוזה עם חלבוני האיתות הקשורים אליהם וממשיכים לאותת מהאנדוזומים או ממדורים תוך-תאיים אחרים. מנגנון זה, מאפשר לליגנד NGF להיקשר ל- RTK הספציפי שלו (הנקרא TrkA) בקצה האקסון הארוך של נוירונים ולאותת לגוף התא שנמצא במרחק רב מקצה האקסון. וזיקולות האיתות האנדוציטיות המכילות TrkA מופעל עם חלבוני איתות המעוגנים לזנב הציטוזולי שלו, מועברים לאורך האקסון אל גוף התא, שם הם מאותתים לתא לשרוד.
שלילת מסיחים:
מסיח ב — נכון. שניהם עוברים אנדוציטוזה פשוט התוצאה הסופית שונה כפי שהוסבר.
מסיח ג — נכון. מוטציות המשביתות את הדאון-רגולציה שעובר הרצפטור ל- EGF או RTKs אחרים של מיטוגנים (למשל מוטציות ב- c-Cbl או בחלבוני UIMs) גורמות לאיתות ממושך של הרצפטור ובכך מקדמות התפתחות סרטן.
מסיח ד — נכון. אם TrkA לא יעבור אנדוציטוזה, הוא לא יצליח לאותת לגוף התא של הנוירון לשרוד ולכן סביר להניח שנראה אפופטוזיס מוגבר.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 913-915
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 25
הפעלת דומיין הקינאז של מי מהרצפטורים הבאים מתבצעת על-ידי דימריזציה וטרנסאוטופוספורילציה?
במה השאלה עוסקת: איתות תאי, RTKs
תשובה ב’. הפעלת הפעילות הקטליטית של רצפטורי טירוזין-קינאז (RTKs) בעת קשירת ליגנד מתבצעת לרוב באמצעות דימריזציה וטרנסאוטופוספרילציה: קשירת ליגנד גורמת לדימריזציה, שפשוט מקרבת בין שני דומייני הקינאז הציטוזוליים, מה שמאפשר להם לזרחן זה את זה על טירוזינים ספציפיים באתרים הפעילים של הקינאז, ובכך לקדם שינויים קונפורמטיביים המפעילים באופן מלא את שני דומייני הקינאז. RTKs רבים, כמו הרצפטור ל- PDGF (ר״ת: platelet-derived growth factor), מופעלים בדרך זו.
שלילת מסיחים:
מסיח א — לא נכון. הוא עובר טרנסאוטופוספורילציה אך לא דימריזציה. הרצפטור לאינסולין קיים כדימר בממברנה גם בהיעדר הליגנד אינסולין, וקשירת הליגנד מכוונת את הדומיינים החוץ-תאיים באופן כזה שמביא את דומייני הקינאז התוך-תאיים לעמדה קרובה יותר זה לזה כדי שיוכלו לזרחן אחד את השני.
מסיח ג — לא נכון. הוא עובר דימריזציה אך לא טרנסאוטופוספורילציה. כאשר EGF נקשר לרצפטור, הוא עובר הדימריזציה שמכוונת את דומייני הקינאז הפנימיים לדימר א-סימטרי, שבו דומיין קינאז אחד
(המכונה activator) דוחף אל עבר דומיין הקינאז השני (המכונה receiver), ובכך גורם לשינוי קונפורמציה משפעל ב- receiver. לאחר מכן, דומיין ה- receiver הפעיל מזרחן מספר שיירי טירוזין בזנבות של שני המונומרים (אין כאן זרחון הדדי).
מסיח ד — לא נכון. אין כזה רצפטור וארסטין הוא לא חלבון סיגנל חוץ-תאי.
משפחת חלבוני הארסטין אלו חלבונים שמעורבים בדה-סנסיטיזציה של GPCRs: הדה-סנסיטיזציה תלויה בזרחון הרצפטור ולאחר מכן הזרחון מזוהה ע״י חלבוני ארסטין שנקשרים ל- GPCR. באופן זה הארסטין יכול למנוע מהרצפטור הפעיל לאקטב חלבוני G, והוא יכול לשמש גם כחלבון מתאם המסייע לאנדוציטוזה תלוית-קלתרין של ה- GPCR.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 911-913
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 26
מי מהבאים גורם לירידה בריכוז של נוקלאוטיד ציקלי בתא?
במה השאלה עוסקת: איתות תאי
תשובה ג’. מערכת האיתות של הראייה נחקרה היטב בתאי rod photoreceptors (קנים) ברשתית של חולייתנים. תאי rods אחראים על ׳ראיית לילה׳ ומנגנון הפוטו-טרנסדוקציה נמצא בסגמנט החיצוני של תא ה- rod, המכיל ערימה של דיסקיות, כאשר ממברנות הדיסקיות עמוסות ב- GPCR הנקרא רודופסין. הממברנה הפלזמטית שמקיפה את הסגמנט החיצוני מכילה תעלות קטיונים–תלויות-cGMP. בתנאי חושך, cGMP הקשור לתעלות הללו שומר אותן פתוחות.
רודופסין קשור קוולנטית לכרומופור שנקרא רטינל, וקליטת פוטון אור (הליגנד) גורמת לאיזומריזציה ברטינל אשר מניעה שינוי קונפרמציה משפעל ברצפטור עצמו, שכעת יכול להפעיל את Gt. תת-יחידה אלפא של Gt מפעילה את האנזים cGMP-פוספודיאסטראז, שבתורו מבצע הידרוליזה של cGMP ובכך מוריד את ריכוזו בציטוזול (וזה גורם לסגירת תעלות הקטיונים ובכך הסיגנל מועבר הלאה).
שלילת מסיחים:
מסיחים א + ב — שני הרעלנים הללו גורמים לעלייה בריכוז ה- cAMP:
רעלן הכולרה מזרז ריבוזילציה של תת-יחידת אלפא של Gs, מה שמונע ממנה לבצע הידרוליזה ל- GTP הקשור. כתוצאה מכל היא נשארת במצב פעיל המגרה את אדנליל-ציקלאז (שמייצר cAMP) ללא הגבלת זמן.
רעלן השעלת מזרז ריבוזילציה של תת-יחידת אלפא של Gi ובכך מונע מהחלבון לקיים אינטראקציה עם ה- GPCR שלו. התוצאה היא שהחלבון G נשאר במצב האינאקטיבי הקושר-GDP ואינו מסוגל לעכב את אדנליל-ציקלאז, מה שיגרום לעלייה בריכוז ה- cAMP.
מסיח ד — כידוע, NO שמיוצר בתאי אנדותל (כאשר אצטילכולין מפעיל GPCR המצומד ל- Gq) מדפייז לתאי השריר החלק של כלי הדם ומפעיל את גואנליל-ציקלאז. העלייה ב- cGMP מובילה להרחבת כלי הדם. התרופה ויאגרה וקרוביה מעכבים את cGMP-פוספודיאסטראז בפין, ובכך מגדילים את משך הזמן שבו רמות ה- cGMP נשארות גבוהות בתאי השריר החלק של כלי הדם בפין לאחר ייצור של NO. ה- cGMP שומר על כלי הדם רפויים ובכך את הפין זקוף.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 895 (רעלן הכולרה והשעלת), 905-906 (איתות במעכ׳ הראייה), 909 (ויאגרה)
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 27
קישור של CO להמוגלובין עלול לגרום למחסור חמור בחמצן ממספר סיבות. איזה מסיח מציג סיבה שגויה?
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה א’. לפחמן חד-חמצני (CO – Carbon monoxide) גודל וצורה הדומים לשל חמצן (O2). עם זאת, CO נקשר הרבה יותר חזק לקבוצת heme חופשית מאשר חמצן (פי 20,000). הכיס החלבוני של המוגלובין אמנם מקטין את הזיקה ל-CO, אבל הוא עדיין נקשר פי 250 יותר טוב מאשר חמצן (שלילת מסיח ד).
ל- CO שנקשר להמוגלובין יש אפקט רעילות כפול:
הוא נקשר לאותו אתר קישור כמו של חמצן, ולכן יהיו פחות אתרים פנויים ב- Hb לקשירת חמצן (שלילת מסיח ג).
הוא מונע שחרור חמצן מכיוון שהוא מאלץ את טטרמר ההמוגלובין להישאר כולו בקונפרמציית R (שלילת מסיח ב).
מסיח א מציג סיבה שגויה — CO אמנם מעלה את זיקת ההמוגלובין לחמצן, אבל זה קורה כי הוא כופה על Hb להיות בקונפרמציית R ולא בקונפרמציית T.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — BOX 5-1
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 28
תאי rods משתמשים במספר מנגנוני משוב שלילי כדי לאפשר לתאים לחזור במהירות למצב מנוחה לאחר הבזק של אור. מי מהבאים מציג מנגנון שגוי?
במה השאלה עוסקת: איתות תאי, איתות דרך Gt
תשובה ד’. כזכור, רודופסין הוא GPCR המצומד ל- Gt (טרנסדוצין). כאשר Gt פעיל הוא מאקטב את cGMP-פוספודיאסטראז, שמוריד את הריכוז של cGMP ובכך נסגרות תעלות קטיונים–תלויות-cGMP. בתנאי חושך, cGMP הקשור לתעלות הללו שומר אותן פתוחות כך שיוני סידן ונתרן נכנסים לתוך התא.
הפסקת האיתות של מעכ׳ הראייה בתאי ה- rod נעשית באמצעות שלושה מנגנונים:
(1) האנזים RK (ר״ת: rhodopsin kinase) מזרחן את הזנב הציטוזולי של רודופסין כך שחלבון ממשפחת הארסטין נקשר לרודופסין המזורחן, מה שמעכב את הרודופסין לשפעל את Gt (וזה למעשה אינאקטיבציה של הרצפטור רודופסין).
(2) חלבון RGS נקשר ל- Gt פעיל ומגרה אותו לבצע הידרוליזה של ה- GTP שלו ל- GDP, מה שמחזיר את Gt למצבו הלא-פעיל (RGS זה הכינוי לחלבוני GAP של חלבוני G הטרוטרימריים).
(3) סגירת תעלות הקטיונים בתגובה לאור גורמת לירידה בריכוז הסידן הציטוזולי. הירידה בריכוז הסידן מעוררת חלבון ספציפי הרגיש ל- +Ca 2 אשר משפעל את גואניליל-ציקלאז (ולכן מסיח ד הוא השגוי). באופן זה ריכוז ה- cGMP חוזר לרמה שהייתה לפני שהאור נדלק ויכול לפתוח בחזרה את התעלות.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 905-907
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני GPCR
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 29
בחקר של חלבון X נמצא שהוא מכיל דומיין SH2. על פי נתון זה, ההנחה הסבירה ביותר היא שהחלבון:
במה השאלה עוסקת: איתות תאי
תשובה ד’. דומיין SH2 בחלבונים מאפשר להקשר לרצף המכיל שיירי טירוזין מזורחנים. רצף של טירוזין מזורחנים נפוץ למצוא במסלול העברת סיגנל של טירוזין קינאז. כחלק מהפעלת הרצפטור חלה טרנסאוטופוספורילציה של החלק התוך תאי של הרצפטור. בעקבות זרחון שיירי הטירוזין, חלבונים בעלי SH2 (המזהים טירוזין מזורחן) יכול להיקשר לרצפטור בחלק התוך תאי ולהוות עוגן לחלבונים נוספים.
שלילת מסיחים:
א. במסלול סיגנל ע”י רצפטור GPCR אין זרחון של שיירי טירוזין ברצפטור ולא נוצר קומפלקס סיגנל על גבי הרצפטור. ברצפטור מסוג זה מופעל חלבון G שבתורו יפעיל אנזים או תעלה.
ב. קינאזות אלו אנזימים אשר מוסיפים קבוצות פוספאט לחלבונים. דומיין SH2 מזהה שיירי טירוזין מזורחנים ולא מבצע את הזרחון בעצמו.
ג. דומיין SH3 מזהה רצפים עשירים בפרולין. נפוץ למצוא חלבונים אשר מכילים גם דומיין SH2 וגם דומיין SH3 (למשל האדפטור Grb2 שמעורב במסלול ההפעלה של קסקדת ה- Ras-MAPK).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 913-916
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מבוא לאיתות תאי
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 30
איזה סרין/תראונין-קינאז מופעל לרוב על-ידי RTK?
במה השאלה עוסקת: איתות תאי
תשובה ב’. הקינאז PKB, שנקרא כך בלנינג׳ר ואילו באלברטס הוא נקרא Akt, מופעל ע״י RTK. הפעלת הקינאז ע״י RTK תלויה בהיקשרות של PI3-קינאז (PI3K, המזרחן פוספואינוזיטידים) לפוספוטירוזין על הרצפטור (או לפוספוטירוזין על IRS1 במקרה של הרצפטור לאינסולין).
הקינאז PKA מופעל ע״י GPCR המצומד ל- Gs.
הקינאז PKC מופעל ע״י GPCR המצומד ל- Gq (שמפעיל את PLC-בטא הגורם להיווצרות IP3, DAG ועלייה בריכוז הסידן הציטוזולי; זכרו שגם RTKs מסוימים מפעילים את PLC-גמא).
הקינאז PKG מופעל ע״י רצפטור גואנליל-ציקלאז (שקף אחרון במצגת ביוכימיה 3 חלק א).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 920-923
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 31
חוקרים ביצעו את שני הניסויים הבאים: בניסוי 1 המוגלובין מאדם בוגר (HbA) הודגר בנוכחות חמצן בלחץ חלקי של 26.8 mmHg, אשר שווה ל-P50 של המוגלובין מהבוגר. בניסוי 2 המוגלובין עוברי (HbF) הודגר בנוכחות חמצן באותו לחץ חלקי של 26.8mmHg. איזו הנחה הכי סבירה?
במה השאלה עוסקת: קשירת חלבונים וליגנדים, המוגלובין
תשובה ב’. תזכורת: Y (או תטא) מתאר את פרקציית החלבון הקושרת ליגנד. ההמוגלובין העוברי HbF הוא בעל אפיניות גבוהה יותר לחמצן מאשר ההמוגלובין של הבוגר, HbA. כלומר, ל-HbF יש P50 נמוך יותר מ-HbA. עובדה זו מאפשרת לעובר להפיק יותר חמצן מהדם שזורם בשלייה ולמנוע תחרות בינו לבין ההמוגלובין של האם. מכיוון ש-HbF הוא בעל אפיניות גבוהה יותר, אם נדגיר אותו ואת HbA באותו לחץ חמצן, HbF יקשור יותר חמצן, כלומר Y שלו יהיה גדול יותר.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Protein-Ligand Interactions Can Be Described Quantitatively
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 32
מה מהבאים אינו תלוי בחלבון IRS1 במסלול האיתות של אינסולין?
במה השאלה עוסקת: איתות תאי, הרצפטור לאינסולין
תשובה א’. הרצפטור לאינסולין (INSR) הוא RTK. כאשר אינסולין נקשר לרצפטור, חלבון המטרה העיקרי שנקשר לפוספוטירוזין על זנב הרצפטור הוא IRS1 (ר״ת: insulin receptor substrate 1). הרצפטור מזרחן שיירי טירוזין גם על IRS1 עצמו, ובדרך זו IRS1 מהווה נקודת הסתעפות שממנה יוצאים שני ענפים עיקריים:
כאשר החלבון המתאם Grb2 (דרך דומיין SH2) נקשר ל- IRS1, מסלול האיתות יוביל לקסקדת ה- MAPK, שדרך מסלול זה אינסולין משפיע על ביטוי הגנים.
תזכורת למסלול: Grb2 -> החלבון Sos (ה- GEF של Ras) -> שפעול Ras -> שפעול Raf, שהוא הקינאז הראשון בקסקדה -> זרחון ושפעול Mek -> זרחון ושפעול Erk. כעת Erk מזרחן מגוון חלבוני מטרה כולל פקטורי שעתוק בגרעין.
כאשר האנזים PI3K נקשר (דרך דומיין SH2) ל- IRS1, יתקבל מסלול האיתות PI3K-PKB (שזה בעצם המסלול שמפעיל את Atk, שנקרא בלנינג׳ר PKB). דרך מסלול זה, אינסולין משפיע על מטבוליזם של גליקוגן והכנסת גלוקוז לתאים. תזכורת למסלול — PI3K מזרחן את PIP2 ליצירת PIP3, מה שמוביל בסופו של דבר לשפעול של PKB. כעת PKB גורם לשני דברים עיקריים:
הוא גורם לאקסוציטוזה של הטרנספורטר GLUT4 לממברנה, ובכך גלוקוז יכנס לתאים.
הוא מזרחן ובכך מעכב את GSK3, מה שמונע ממנו לעכב את גליקוגן-סינתאז וכך תאי שריר ושומן יסנתזו גליקוגן מכל הגלוקוז שנכנס.
אפקסין לא קשור למסלול של אינסולין, אלא הוא ה- GEF של Rho והוא מעורב במסלול של RTKs ממשפחת Eph (שגורמים לקריסת חרוט הגדילה של אקסון נודד כדי לכוון אותו לשריר המטרה שהוא אמור לעצבב).
להרחבה — Lehninger מהדורה שמינית, פרק 12.4
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 33
העקומה מתארת את הפרקציה הקושרת ליגנד של חלבון X (משמע ערך Y) כתלות בריכוז הליגנד שלו. בהסתמך על עקומה זו, מבין ההיגדים הבאים – מי הכי סביר בנוגע לחלבון X?
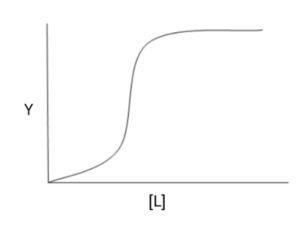
במה השאלה עוסקת: קשירת חלבונים וליגנדים
תשובה ג’. הצורה של עקומת הקישור היא סיגמואידלית, מה שמרמז לנו על כך שחלבון X קושר את הליגנד שלו באופן קואופרטיבי (= שיתופי. שיתופיות חיובית = קשירת ליגנד מעלה את האפיניות של שאר אתרי הקישור לליגנד, ואילו שיתופיות שלילית = קישור ליגנד מוריד את האפיניות של שאר האתרים).
חלבונים המדגימים קישור קואופרטיבי הם חלבונים אלוסטרים. אם הרגולטור האלוסטרי זהה לליגנד הטבעי של החלבון אז הוא נקרא מודולטור הומוטרופי ואם הוא שונה אז הוא מודולטור הטרוטרופי.
קבוע היל מתאר את מידת הקואופרטיביות של החלבון: אם הקבוע גדול מ-1 הקישור הוא קואופרטיבי חיובי, אם הוא קטן מ-1 הקישור הוא קואופרטיבי שלילי, ואם הוא שווה ל-1 הקישור אינו קואופרטיבי (מה שמאופיין בעקומת קישור היפרבולית).
הניסוח של מסיח ג׳ לא הכי מדויק ומנסה לבלבל אבל הוא המסיח הנכון ביותר משתי סיבות:
כל שאר המסיחים ניתנים לשלילה בקלות (פירוט למטה).
השיתופיות החיובית מתבטאת בגרף כאשר ריכוז הליגנד מעט עולה והחלבון מתחיל לקשור ליגנד, כך ששאר אתרי הקישור הריקים לאותו ליגנד עוברים מאפיניות נמוכה לאפיניות גבוהה, ולכן במצב בו האתרי קישור עם אפיניות גבוהה — ערך ה- Kd בפועל אמור לרדת.
שלילת מסיחים:
א. על מנת שתתקבל עקומה סיגמואידלית שמעידה על קישור קואופרטיבי צריכים להיות לפחות שני אתרי קישור לליגנד.
ב. המודלטור האלוסטרי במקרה זה הוא הומוטרופי, משמע הוא גם הליגנד הטבעי של החלבון.
ד. קבוע היל במקרה זה גדול מ-1 מכיוון שמוצגת עקומת קישור עם קואופרטיביות חיובית: רואים שהחל מריכוז ליגנד מסוים — העקומה עולה בחדות כך שהחלבון קושר במהירות עוד ליגנד עד הגעה לריוויון. אם החלבון היה מקיים שיתפיות שלילית, אז העקומה אולי הייתה סיגמאודית אבל אין שום סיכוי שהיא הייתה נראת כך, וכמובן שלא היינו רואים עלייה חדה בעקומה כאשר החלבון מתחיל לקשור ליגנד.
כמעט ואין דוגמאות אמיתיות לקואופרטיביות שלילית בטבע, ולכן לא בדיוק מובן איך תראה עקומת הקישור של חלבון היפותטי המקיים שיתופיות שלילית (אם הייתי צריכה לנחש אז אולי זה היה נראה גרף סיגמואידי הפוך, כלומר ירידה בקישור החל מריכוז ליגנד מסוים ולא עלייה). בכל אופן מכיוון שזה נדיר בטבע ולא מדובר בספר — אין סיבה שתשאלו על עקומת קישור עם שיתופיות שלילית. המסיח הזה נמצא כאן כדי שלא רק תדעו לשנן שעקומה סיגמאודית משקפת שיתופיות, אלא שתדעו להבין כיצד מתבטאת השיתופיות החיובית בגרף.
להרחבה — Lehninger מהדורה שמינית, פרק 5.1
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 34
מי מהבאים נכון בנוגע למסלול האיתות Wnt/בטא-קטנין?
במה השאלה עוסקת: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ד’. חלבוני Wnt הם מולקולות סיגנל מופרשות השולטות בהיבטים שונים של התפתחות בעלי חיים. בטא-קטנין הוא חלבון תוך-תאי ויש לו שתי פונקציות שונות:
חלק מהבטא-קטנין בתא משתתפים בצמתי תא-תא כחלבונים מתאמים (כפי שנראה בביותא 4).
שאר הבטא-קטנין שלא משתתפים בצמתים, מתפקדים כבקרי שעתוק לטנטיים. מסלול האיתות של Wnt פועל בעיקר עליהם.
תיאור המסלול (ראו איור):
(A) בהיעדר הסיגנל Wnt, הבטא-קטנין שאינו מצוי בצמתים מקיים אינטראקציה עם קומפלקס דגרגציה המכיל את חלבוני הפיגום APC ו- axin ואת הקינאזות CK1 ו- GSK3, אשר מזרחנים אחד אחרי השני את בטא-קטנין, מה שגורם ליוביקוויטילציה ולדגרגציה שלו בפרוטאזומים. במצב זה, גני המטרה של Wnt מעוכבים ע״י קו-רפרסור בשם Groucho.
(B) קישור Wnt ל- Frizzled ול- LRP מפגיש בין שני הקו-רצפטורים הללו, והזנב הציטוזולי של LRP מזורחן ע״י CK1 ו- GSK3.
חלבון הפיגום Dishevelled מגויס לחלבון Frizzled הפעיל. כעת axin נקשר ל- Dishevelled ול- LRP המזורחן, ובכך axin מושבת וקומפלקס הדגרגציה מתפרק.
בדרך זו נמנע הזרחון של בטא-קטנין, כך שהוא יכול להיכנס לגרעין, שם הוא נקשר לבקר השעתוק LEF1/TCF, מחליף את הקו-רפרסור Groucho, ופועל כקו-אקטיבטור כדי לעורר את שעתוק גני המטרה של Wnt.
חלק מהגנים שבטא-קטנין מעורר מדכאים את סיגנל ה- Wnt והתוצאה היא משוב השלילי, לדוגמה: אחד מהגנים הללו מקודד לחלבון מופרש שמסיר את המודיפיקצייה הליפידית מ- Wnt ובכך משבית אותו.
שלילת מסיחים:
מסיח א — Wnt הוא חלבון ולא ליפיד. חלבוני Wnt מחוברים קוולנטית לחומצות שומן, ולכן הם נחשבים די הידרופוביים ונוטים להתחבר לממברנות התאים במקום לדפייז במהירות בסביבה החוץ-תאית. מסיבה זו מניחים שהם פועלים בעיקר באופן פאראקריני (מקומי). המודיפיקציה הליפידית של Wnt הכרחית לפעילותו (כי הקו-רצפטור Frizzled מכיל כיס הידרופובי וכך Wnt נקשר אליו בזיקה גבוהה).
מסיח ב — המסיח הזה היה נכון אם היה כתוב APC במקום Myc. חלבון הפיגום APC (ר״ת: Adenomatous polyposis coli) מוטנט ברוב מקרי סרטן המעי הגס. המוטציות מעכבות את יכולתו של APC לקשור את בטא-קטנין בקומפלקס הדגרגציה, כך שהוא נכנס לגרעין גם בהיעדר Wnt ומפעיל את הגנים התלויים בו (ביניהם הפרוטואונקוגן Myc שמעודד את התא להתחלק, כפי שלמדתם בביותא 2).
מסיח ג — להפך, הוא גורם להתפרקות קומפלקס הדגרגציה ושחרור של בטא-קטנין.
מסיח ה — כאמור, APC הוא חלבון פיגום בקומפלקס הדגרגציה.
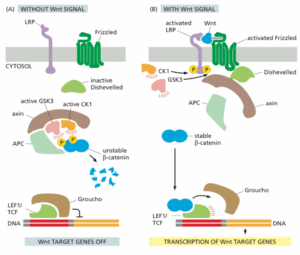
להרחבה – Alberts, מהדורה שביעית, עמ׳ 930-931
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 35
חלבוני Hedgehog וחלבוני Wnt פועלים בדרכים דומות. מי מהבאים לא מציג נקודת דימיון ביניהם?
במה השאלה עוסקת: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה : א. בשונה מחלבוני Wnt, חלבוני Hedgehog הם חלבוני סיגנל שיוזמים מסלול איתות מורכב בסיליה הראשונית (primary cilium). ראו איור:
(A) בהיעדר הליגנד Hedgehog, הרצפטור שלו (שנקרא Patched) שוכן בממברנת הסיליום ומעכב את הפעילות של חלבון טרנסממברנלי אחר בשם Smoothened, שנמצא מחוץ לסיליום. חושבים שהעיכוב נעשה ע״י כך ש- Patched מפחית את כמות הכולסטרול בממברנת הסיליום.
גני המטרה של Hedgehog נשמרים לא-פעילים בשתי דרכים:
ראשית, בקרי השעתוק Gli (שמתפקדים כאקטיבטורים של ביטוי הגנים) מוחזקים במצב לא-פעיל ע״י SuFu.
שנית, רצפטור GPCR פעיל (Gpr161) מאקטב את אדניליל-ציקלז לסנתז cAMP, מה שמוביל לזרחון תלוי-PKA של החלבון Gli3. במצבו המזורחן, Gli3 מעובד ליצירת רפרסור שעתוק שמעכב את ביטוי הגנים.
(B) קשירת Hedgehog לרצפטור Patched גורמת לשני דברים:
הסרת העיכוב מ- Smoothened (כנראה כי קישור הליגנד ל- Patched חוסה את ערוץ הובלת הכולסטרול שלו) כך שהוא עובר לממברנת הסיליום, שם הוא גורם לניתוק SuFu מ- Gli2 והם כעת יכולים להיכנס לגרעין ולעורר ביטוי של גנים המגיבים ל- Hedgehog.
קישור Hedgehog לרצפטור מקדם הסרה של Gpr161 מהסיליום (לא מוצג), ובכך מפחית את העיבוד של Gli3 לרפרסור שעתוק.
האיתות של Hedgehog יכול לקדם פרוליפרציה של תאים, כך שאיתות יתר של Hedgehog יכול להוביל לסרטן.

להרחבה – Alberts, מהדורה שביעית, עמ׳ 932-933
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 36
מה מהבאים נכון בנוגע לאיתות בצמחים?
במה השאלה עוסקת: איתות תאי, איתות בצמחים (מצגת ביוכימיה 3 חלק ב׳)
תשובה א’. בצמחים, כמו בבעלי חיים, התאים נמצאים בתקשורת מתמדת אחד עם השני.
רכיבי איתות כמו cGMP ,יוני סידן, NO, פרוטאין-קינאזות ו- GTPases מונומריים נמצאים גם בצמחים וגם בבע״ח.
צמחים לא מכילים הומולוגים ממשפחת הרצפטורים הגרעיניים, וגם לא הומולוגים של Wnt ,Notch ,TGFβ ,STAT ,JAK ,Hedgehogבאופן דומה, הם גם לא משתמשים ב- cAMP לאיתות תוך-תאי.
האסטרטגיות הכלליות העומדות בבסיס האיתות דומות בצמחים ובבע״ח, למשל שניהם משתמשים ברצפטורי cell-surface המצומדים לאנזים. עם זאת, בעוד שקולטני RTK הם הסוג הנפוץ של רצפטור המצומד לאנזים בבע״ח, בצמחים RTKs הם נדירים ביותר והמשפחה הגדולה ביותר של רצפטורים בפני שטח התא אלו קולטני סרין/תראונין-קינאז.
שלילת מסיחים:
מסיח ב — לצמחים אין Notch (וזה הגיוני כי אין להם תאי אפיתל ונוירונים).
מסיח ג — ההורמון הצמחי אתילן עושה את זה.
מסיח ד — פוטופרוטאינים (photoprotein) הם קבוצת חלבונים רגישים לאור שנמצאת בצמחים. כל הפוטופרוטאינים חשים אור באמצעות מולקולת כרומופור אשר קשורה אליהם באופן קוולנטי. הכרומופור משנה את צורתו בתגובה לאור ולאחר מכן גורם לשינוי בקונפורמציה של החלבון.
מסיח ה — הם לא משתמשים ב- cAMP.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 940-945
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 37
מי מהבאים הכי פחות סביר שהוא שייך למשפחת הרצפטורים הגרעיניים (nuclear receptor superfamily)?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ד׳. מספר מולקולות סיגנל קטנות והידרופוביות, מדפייזות ישירות דרך הממברנה הפלזמטית של תאי המטרה ונקשרות לרצפטורים תוך-תאיים הפועלים כבקרי שעתוק. מולקולות איתות חוץ-תאיות אלה כוללות הורמונים סטרואידים, הורמוני בלוטת התריס, רטינואידים וויטמין D. כל הליגנדים הללו פועלים באמצעות מנגנון דומה: הם נקשרים לרצפטור החלבוני התוך-תאי שלהם ומשנים את היכולת של חלבונים אלה לשלוט בשעתוק של גנים ספציפיים. כל הרצפטורים הגרעיניים נקשרים לדנ”א כהומודימרים או כהטרודימרים, ולכן מסיח ד’ הוא השגוי.
שלילת מסיחים:
מסיח א — נכון. הליגנדים של חלק מהרצפטורים הגרעיניים ביונקים הם מטבוליטים תוך-תאיים ולא מולקולות סיגנל מופרשות. למשל PPARs קושרים מטבוליטים תוך-תאיים של ליפידים ומבקרים את השעתוק של הגנים המעורבים במטבוליזם של ליפידים ובהתמיינות תאי שומן.
מסיח ב — נכון, כפי שמוסבר למעלה.
מסיח ג — נכון. חלק מהרצפטורים הגרעיניים (כמו הרצפטור לקורטיזול) ממוקמים בציטוזול ונכנסים לגרעין רק לאחר קשירת ליגנד . רצפטורים גרעיניים אחרים קשורים לדנ”א בתוך הגרעין גם בהיעדר ליגנד (אך הוא לא יהיה פעיל עד שהליגנד יקשר אליו).
מסיח ה — נכון. לעתים קשירת הליגנד לרצפטור גורמת לו לקשור קו-אקטיבטורים שמעוררים שעתוק גנים, אך במקרים אחרים, קשירת הליגנד לרצפטור מעכבת שעתוק: למשל חלק מהרצפטורים להורמוני בלוטת התריס, פועלים כאקטיבטורים של שעתוק בהיעדר ההורמון שלהם והופכים לרפרסורים של שעתוק כאשר ההורמון נקשר אליהם.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 935-937
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 38
מי מהבאים אחראי על אקטוב תהליך העיכוב הלטרלי (lateral inhibition)?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה א׳. איתות דרך החלבון Notch נמצא בשימוש נרחב במהלך ההתפתחות של בעלי חיים. הוא ידוע בעיקר בשל תפקידו בייצור תאי עצב בדרוזופילה, אשר בדר״כ מופיעים כמספר תאים מבודדים בתוך יריעת אפיתל של תאי פרקורסור. במהלך תהליך זה, כאשר תא פרקורסור מתחייב להפוך לתא עצבי, הוא מאותת לתאים השכנים לא לעשות את אותו הדבר (התאים המעוכבים מתפתחים לתאי אפידרמיס במקום).
תהליך זה נקרא עיכוב לטראלי (lateral inhibition) והוא תלוי במנגנון איתות תלוי-מגע המופעל ע״י דלתא, שהוא חלבון סיגנל טרנסממברנלי החוצה את הממברנה פעם אחת, ומצוי בפני השטח של תא העצב העתידי (ולכן מסיח א הוא הנכון). על-ידי קשירה של דלתא לרצפטור שלו (החלבון Notch) על התא השכן, דלתא מאותת לתא השכן לא להפוך לתא עצב. כאשר תהליך האיתות הזה פגום, נוצר עודף עצום של תאי עצב על חשבון תאי האפידרמיס, וזה כמובן מצב קטלני שלא מאפשר חיים.
ברוב המקרים, כל התאים בצביר מבטאים בהתחלה גם את דלתא וגם את Notch, כך שמתרחשת תחרות כאשר תא אחד מסתמן כמנצח – התא שמבטא את דלתא בחוזקה ובכך מונע משכניו לעשות כמוהו, הוא ה״מנצח״.
שלילת מסיחים:
מסיח ב — גמא-סקרטאז הינו פרוטאז שמעורב בעיבוד הפרוטאוליטי של Notch כחלק ממסלול האיתות.
מסיח ג — כמו בכל מסלולי האיתות, מי שמאקטב את התהליך הוא מולקוםת הסיגנל הנקשרת לרצפטור (הליגנד), והליגנד הוא כמובן דלתא. שימו לב גם שהתהליך מתרחש כאשר דלתא על פני תא העצב העתידי נקשר לרצפטור Notch על פני השטח תאי הפרקורסור שצריכים להיות מעוכבים.
מסיח ד — דלתא הוא חלבון סיגנל טרנסממברנלי (ולכן מדובר באיתות תלוי-מגע) ואינו חלבון סיגנל מופרש.

להרחבה – Alberts, מהדורה שביעית, עמ׳ 928-930
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 39
חברתכם הטובה התחתנה עם מדען מטורף שמקדיש את חייו לבניית שעון צירקדי. החברה פלטה בתום לב שאתם לומדים למבחן הידע ומאז המדען משגע אתכם לעזור לו… לפניכם כמה מהגישות של המדען ביחס לבניית השעון. איזו גישה כן תמליצו לו להשאיר בתוכנית הבנייה כדי שיגשים את חלומו במהרה ויעזוב אותכם לנפשכם?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ד׳. שאלה לא חביבה שמטרתה להעביר את העקרונות הכלליים של שעונים צירקדיים. ראשית, ננסה להסביר את העקרון (אין פה העמסה של פרטים לשינון):
לאורגניזמים רבים יש מיקצב פנימי המווסת התנהגויות שונות בשעות שונות של היום והלילה. הרכיבים התוך-תאיים השולטים במקצבים יומיים כאלה מכונים אוסילטורים (מתנדים) והם בעצם מרכיבים את השעון הצירקדי. כל המטרה של שעון צירקדי זה לשלוט בביטוי הגנים בדפוס מחזורי של 24 שעות, כלומר במהלך היום יתבטאו לנו גנים מסוימים שנדרשים ליום ואילו במהלך הלילה דפוס הביטוי ישתנה כדי לבטא גנים שאנו זקוקים להם בלילה. כדי שנקבל תנודות של 24 שעות בביטוי הגנים, נדרש משוב שלילי בדיליי (כמו שראינו עם התנודתיות בריכוז הסידן הציטוזולי, פשוט כאן כל תנודה חוזרת על עצמה בפרק זמן של 24 שעות).
למשל גן A מתחיל להתבטא בשמונה בבוקר ומתחיל לדעוך משעות הערב, ואז למחרת הוא שוב מתבטא בשמונה בבוקר וכן הלאה — אם נצייר את הביטוי של גן A בגרף (כאשר ציר x מייצג זמן ולא את השעה ביום), זה יראה בערך ככה:
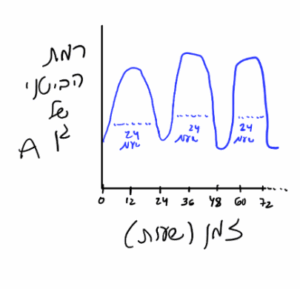
כדי ששעון צירקדי יועיל לאורגניזם (כלומר השעון יהיה מסונכרן עם הסביבה), הוא צריך להיות מסוגל להתאפס ע״י אותות מהסביבה שמציינים את השעה ביום (שזה בעיקר אור וחושך). למשל אם נטוס פתאום לארה״ב — אז גן A יתבטא בלילה ביום-יומיים הראשונים, אך השעון יתאפס בהדרגה כדי שהתנודות של ה- 24 שעות יסתדרו כך שגן A יתחיל להתבטא בשמונה בבוקר לפי שעון ארה״ב.
בחזרה לשאלה: מסיח ד מתאר את עיקרון הליבה של השעון — כדי שהיו תנודות של 24 שעות, צריך שהיה משוב שלילי בדיליי. הספר מציג שתי דוגמאות למנגנון של שעון צירקדי: אחת בדרזופילה ואחת בציאנובקטריה. שניהם מבוססים על משוב שלילי בדיליי, אך המנגנון כמובן שונה.
ביונקים, תאי ה- SCN הם הרגולטורים המרכזיים של המקצב הצירקדי:
תאי ה- SCN מקבלים אותות עצביים מהרשתית וכך הם נכנסים למחזור היומי של אור וחושך. תאי ה- SCN שולחים מידע על השעה ביום לאזורים אחרים במוח, שבתורם מעבירים את סיגנל הזמן הזה לשאר הגוף על-ידי שחרור ההורמון מלטונין בזמן לפי השעון.
חשוב להבין שהשעון הצירקדי פועל בכל אחד מתאי ה- SCN: גם אם התאים הללו יוסרו מהמוח ויפוזרו בצלחת תרבית, הם ימשיכו להתנודד בנפרד ולהראות דפוס מחזורי של ביטוי גנים עם פרק זמן של כ- 24 שעות.
לרוב התאים האחרים בגופנו יש גם שעונים צירקדיים, שיש להם את היכולת להתאפס בתגובה לאור.
הספר לא מפרט על המנגנון של ה- SCN והשורה התחתונה היא ששעונים צירקדיים הם תכונה בסיסית של תאים רבים (אין פה איזשהו מנגנון רב-תאי מורכב).
שלילת מסיחים:
מסיח א — חבל מאוד שהמדען לא אוהב ציאנובקטריות כי הוא היה יכול לחסוך המון כאב ראש. השעון הצירקדי המובן ביותר נמצא בציאנובקטריה והוא מורכב משלושה חלבונים בלבד ( KaiA, KaiB ו- KaiC). אם נבודד את שלושת החלבונים הללו במבחנה נקבל שעון צירקדי: השחקן המרכזי הוא KaiC, אנזים מולטי-פונקציונלי שמזרז את הזרחון והדה-זרחון של עצמו במחזוריות של 24 שעות. בציאנובקטריה עצמה, מצב הזרחון של KaiC משפיע בעקיפין על בקרי שעתוק ובכך על ביטוי הגנים.
מסיח ב — כפי שהוסבר למעלה, הניסיון של המדען להתמקד בבניית שעון שיכלול מיליון תאים מסונכרנים זה מיותר ולא ריאלי, מכיוון שלתאים רבים (כולל כל אחד מתאי ה- SCN) יש שעון צירקדי משל עצמו אשר יכול ליצור תנודות של 24 שעות ללא עזרה מתאים אחרים.
מסיח ג — מיותר. הסימנים מהסביבה של אור וחושך זה כדי לסנכרן את המקצב הצירקדי של האורגניזם עם סביבתו. אם השעון לא יקבל שום סימן מהסביבה (למשל יהיה בחושך מוחלט) — השעון עדיין ימשיך לרוץ ולהראות תנודות של 24 שעות פלוס מינוס.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 937-939
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 40
התגלה עובר דרזופילה מוטנט שאינו שרד את ההתפתחות. החוקרים הבחינו שהעובר מכיל עודף של תאי עצב על חשבון תאי אפיתל והסיקו שהוא נשא פגם במסלול העיכוב הלטראלי, אם כי הם לא הצליחו לאתר היכן בדיוק המוטציה. מי מההשערות הבאות הכי פחות סבירה למקור המוטציה?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ב׳. כדי לענות על השאלה, יש לחשוב כיצד כל מוטציה יכולה לפגוע בתהליך האיתות התקין כך שבתא הפרקורסור לא יופעל השעתוק של ה- Notch-response genes (ואז הוא עלול להפוך לתא עצב). תחילה יש להכיר את שלושת שלבי העיבוד הפרוטאוליטיים שעובר Notch (ראו איור):
(1) שלב העיבוד הראשון מתרחש בגולג׳י כדי ליצור את הרצפטור הבוגר — Notch הטרודימרי, שיועבר לאחר מכן לממברנת התא.
שני השלבים הפרוטאוליטיים הבאים מתרחשים כאשר Notch נקשר לדלתא על פני התא הסמוך:
(2) הקומפלקס של דלתא ומקטע ה- Notch שאליו הוא נקשר עוברים אנדוציטוזה לתוך התא שמבטא את דלתא, מה שחושף את אתר הביקוע החוץ-תאי בתת-היחידה הטרנסממברנלית של Notch (הביקוע נעשה ע״י פרוטאז חוץ-תאי).
(3) כעת מתרחש הביקוע השלישי (ע״י גמא-סקרטאז) כדי לשחרר את זנב ה- Notch מהממברנה.
הזנב המשוחרר נודד לתוך הגרעין (של התא המיועד להיות תא אפיתל) שם הוא נקשר לחלבון Rbpsuh, ובכך ממיר אותו מרפרסור שעתוק לאקטיבטור שעתוק. באופן זה זנב ה- Notch מפעיל את שעתוק הגנים הרלוונטים (Notch-response genes) ובכך התא יהפוך לתא אפידרמלי ולא לתא עצב.
מסיח ב מציג השערה שגויה מכיוון שאם Rbpsuh יתפקד כאקטיבטור גם ללא קישור של הזנב — אז הסט גנים יתבטא גם כאשר דלתא לא נקשר ל- Notch ובמצב זה דווקא עלול להיות מחסור בתאי עצב (כי Rbpsuh יתפקד כאקטיבטור בכל התאים ברקמה המתפתחת).
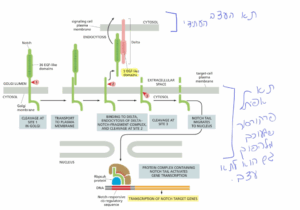
שלילת מסיחים:
מסיח א — השערה סבירה. אם Notch לא יעבור עיבוד תקין בגולג׳י לא יהיה רצפטור Notch בוגר (הטרודימר) תקין בממברנה וכנראה שכל המסלול ייכשל.
מסיח ג — השערה סבירה. לאחר שדלתא נקשר ל- Notch מתרחשת אנדוציטוזה של דלתא ומקטע הנוץ׳ לתוך תא העצב העתידי, מה שחושף את אתר הביקוע השני במקטע הנוץ׳ שנשאר בממברנת תא הפרקורסור, ולכן חוסר פעילות של החלבון האנדוציטי בתא העצב העתידי יגרום לכישלון העיכוב הלטראלי.
מסיח ד — השערה סבירה. כדי שהמסלול יתפקד נדרש Notch בוגר ותקין על הממברנה, ופגם בגליקוזילציה שלו עלול למנוע זאת.
מסיח ה — השערה סבירה. האינטראקציה בין דלתא לבין המקטע החיצוני של Notch נעשית דרך הדומיינים דמויי-EGF החוזרניים שלהם, ולכן מוטציה בהם עלולה לפגוע בקישור בין הרצפטור לליגנד וכישלון העיכוב הלטראלי.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 928-930
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 41
סמנו את המשפט הנכון
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ג׳. מסלול האיתות של Hedgehog מתואר בהרחבה בהסבר לשאלה 35, והמסלול של Wnt בשאלה 34.
איתות יתר של Wnt עלול להוביל לסרטן (בעיקר סרטן המעי הגס), בין היתר כי החלבון Myc יתבטא יתר על המידה במצב כזה ויוביל לפרוליפרציה בלתי-מבוקרת של התאים. החלבון APC (ר״ת: Adenomatous polyposis coli) מעורב במסלול האיתות של Wnt, שם הוא מהווה חלק מקומפלקס הדגרגציה אשר גורם לפירוק בקר השעתוק הרדום בטא-קטנין (ובכך גני המטרה של Wnt נשמרים לא-פעילים).
השם של APC ניתן לו כי גילו שהוא לעתים קרובות מוטנט בסוג של גידול שפיר (אדנומה) במעי הגס. הגידול מקרין לתוך לומן המעי כפוליפ ובסופו של דבר יכול להפוך לממאיר, ולכן זה לא מפתיע שמוטציות בגן המקודד ל- APC מתרחשות ב- 80% ממקרי סרטן.
גם איתות יתר של Hedgehog יכול לקדם פרוליפרציה של תאים ובכך סרטן, לדוגמה — מוטציה משביתה ב- Patched, שמובילה לאיתות יתר של Hedgehog, מתרחשות לעתים קרובות ב- basal cell carcinoma של העור. זה קצת מבלבל מכיוון שבדר״כ נצפה שאם מוטציה משביתה רצפטור איתות, אז זה יוביל לחסר במסלול האיתות ולא לאיתות יתר. עם זאת, באיתות של Hedgehog קורה משהו קצת שונה: קישור של Hedgehog לרצפטור Patched גורם לעיכוב הרצפטור ולא לשפעול שלו, ובכך העיכוב של Smoothened מתבטל.
מולקולה קטנה בשם ציקלופמין משמשת לטיפול בסרטן הקשור לאיתות יתר של Hedgehog:
ציקלופמין חוסם את האיתות של Hedgehog ע״י קישור הדוק ל- Smoothened ועיכוב פעילותו. באופן זה המולקולה הזו משמשת לטיפול בסרטן.
הציקלופמין זוהה במקור משום שהוא גרם לפגמים התפתחותיים חמורים בצאצאי כבשים שרעו באזור המכיל את הפרחים שמייצרים את המולקולה הזו. הפגמים כוללים נוכחות של עין מרכזית אחת (מצב הנקרא ציקלופיה), מצב שנראה גם בעכברים הלוקים בחסר של Hedgehog signaling.
שלילת מסיחים:
מסיח א — ציקלופמין משמשת לטיפול בסרטן ע״י קישור והשבתה של Smoothened (אשר מעוכב ע״י Patched בהיעדר Hedgehog).
מסיח ב — ציקלופה היא תוצאה של מחסור באיתות של Hedgehog, עקב כך שהכבשים בהריון אכלו את הפרח שמייצר ציקלופמין והעוברים שלהם שילמו את המחיר.
מסיח ד — זה נפוץ כתוצאה מאיתות יתר של Hedgehog.
מסיח ה — מוטציות משביתות כאלה גורמות לאיתות יתר של Hedgehog כפי שמוסבר למעלה.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 930-933
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 42

במה השאלה עוסקת: קשירת חלבונים וליגנדים
תשובה ב’. ליגנד נקשר לחלבון בריאקציה שלה יש קבוע קצב (ka) מסויים, ומשתחרר מהחלבון בריאקציה עם קבוע קצב אחר (kd) היחס בין ka/kd למעשה מתאר את קבוע שיווי המשקל K (שהוא קבוע האסוציאציה Ka) והיחס ההפוך מתאר את קבוע הדיסוציאציה Kd. על מנת לענות על השאלה יש למצוא את Kd מכיוון שהוא מייצג את ריכוז הליגנד שבו נקבל Y=0.5 (משמע 50% מסך החלבון קושר ליגנד).
בריאקציה שבה ליגנד נקשר לחלבון עם קבוע קצב ka ומתפרק ממנו עם קבוע קצב kd, קבוע הדיסוציאציה שהוא מדד הופכי לאפיניות בין הליגנד לחלבון שווה ליחס בין קבוע הקצב kd ל- ka:

להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Protein-Ligand Interactions Can Be Described Quantitatively
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 43
הנוגדן המונוקלונאלי Adalimumab נקשר לליגנד שלו TNF-α עם Kd של X מולר. איזה ריכוז של TNF- α נדרש על-מנת ש-80% מסך כל אתרי הקישור של Adalimumab במבחנה יהיו תפוסים?
במה השאלה עוסקת: קשירת חלבונים וליגנדים
תשובה ב’. נזכיר את הנוסחא לחישוב Y – הפרקציה הקושרת אתרי הקישור בחלבון שתפוסים ע”י הליגנד:
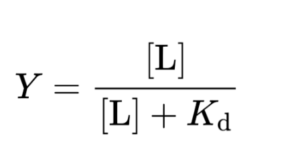
כאשר: [L] הוא ריכוז הליגנד ו-Kd הוא קבוע הדיסוציאציה. Kd הוא ריכוז הליגנד שבו Y שווה 50%. אם נציב ב-Y את הערך 0.8 כמו בשאלה ובמקום Kd נציב X ונחלץ את ריכוז הליגנד נקבל שריכוז הליגנד (הנדרש כדי ש- 80% מסך אתרי הקישור יהיו תפוסים) הוא 4X.
אופן הפישוט המתמטי למעוניינים:
![]()
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — Protein-Ligand Interactions Can Be Described Quantitatively
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 44
טווח ה- Kd של מי מהצמדים הבאים הוא הנמוך ביותר?
במה השאלה עוסקת: קשירת חלבונים וליגנדים
תשובה ב’. זוהי שאלה שדורשת מכם להכיר בצורה טובה את הקישור של כל צמד. עם זאת, אפשר לנסות לענות על השאלה גם מבלי לזכור: אפשר לבחון האם נדרשת אינטראקציה חזקה (ולכן Kd נמוך) בין כל צמד או שלא…
רצפטור-ליגנד, ה- Kd שלהם לא יהיה הנמוך ביותר, מאחר ולצמדים רבים של רצפטור-ליגנד צריכה להיות יכולת להתנתק בצורה יחסית קלה כדי לבצע את תפקודם הביולוגי (למשל המוגלובין צריך לדעת גם לשחרר חמצן ולא רק לקשור אותו).
רצף דנ”א-חלבון, לרוב יהיו אלו פקטורי שעתוק שייקשרו לדנ”א, או רפרסורים. אלו חלבונים שמתנתקים ומתחברים בצורה די חזקה, אך עדיין, נצפה מהם להיות בעלי יכולת להתנתק כשצריך.
אנזים-סובסטרט, אמנם הספציפיות ביניהם גבוהה, אך כל תפקידו של אנזים הוא לזרז ריאקציה ולא לקשור בחוזקה את הסובסטרט, ולכן ה- Kd שלהם לא יהיה גבוה.
נותרנו עם נוגדן-אנטיגן. על מנת שנוגדן יוכל לבצע את תפקידו ביעילות רבה, עליו להיות מאוד ספציפי לאנטיגן, ולא לשחרר אותו (לא היינו רוצים שפתוגן מסוים שנקשר, יתנתק מהנוגדן לאחר זיהוי).
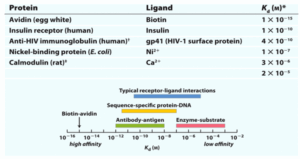
להרחבה — Lehninger מהדורה שמינית, פרק 5.1 — TABLE 5-1 Protein Dissociation Constants: Some Examples and Range
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – חלבונים קושרי-חמצן ואלוסטריה
ספר – Lehninger 8th edition
פרק – 5
ביוכימיה 4 שאלה 45
באיזה מסלול איתות הסיגנל החוץ-תאי לא מפעיל מעבר מדיכוי השעתוק לאקטיבציית השעתוק?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה א’. המסלול של Hedgehog, של Wnt ושל Delta (עיכוב לטראלי) מתוארים בהרחבה בשאלות 35, 34 ו- 40 (בהתאמה).
חלבוני NFkB הם בקרי שעתוק לטנטים הנמצאים ברוב תאי בעלי החיים והם מרכזיים בתגובות דלקתיות וחיסוניות רבות. רצפטורי cell-surface שונים מפעילים את מסלול האיתות של NFkB בתאי בעלי חיים: לדוגמה בחולייתנים, Toll-like receptors מזהים פתוגנים ומפעילים את המסלול הזה כדי לעורר את תגובת מעכ׳ החיסון, וגם הרצפטורים עבור הציטוקינים הדלקתיים TNF-אלפא ו- IL1 מפעילים את מסלול האיתות הזה.
בהיעדר סיגנל חלבוני NFkB מוחזקים ע״י קומפלקס חלבונים מעכב, מה שמונע את הביטוי של הגנים הרלוונטים מכיוון שהשעתוק שלהם דורש שחלבוני NFkB יקשרו אליהם (בניגוד לשלושת המסלולים האחרים, שבהיעדר סיגנל – עיכוב שעתוק הגנים מערב בין היתר נוכחות של רפרסורים שדואגים לדכא את השעתוק).
כאשר המסלול מופעל (למשל ע״י קשירת TNF-אלפא לרצפטור שלו), העיכוב על NFkB מתבטל והם נכנסים לגרעין ומעודדים את שעתוק הגנים הרלוונטים.
שלילת מסיחים:
מסיח ב — לאחר העיבודים הפרוטאולטיים – הזנב של Notch נקשר לבקר השעתוק Rbpsuh וממיר אותו מרפרסור שעתוק לאקטיבטור שעתוק.
מסיח ג — לאחר פירוק קומפלקס הדגרגציה – בטא-קטנין מצטבר ונכנס לגרעין, שם הוא נקשר ל- LEF1/TCF, מחליף את הקו-רפרסור Groucho ופועל כקו-אקטיבטור כדי לעורר את שעתוק גני המטרה של Wnt.
מסיח ד — בהיעדר סיגנל, Gli3 עובר ביקוע פרוטאוליטי כדי ליצור רפרסור שעתוק ששומר על גני המטרה של Hedgehog מושתקים. בנוסף, הרצפטור (Patched) חוסם את ההפעלה של Gli2. לקישור של Hedgehog ל- Patched יש שתי השפעות: הקישור מבטל את העיבוד הפרוטאוליטי של Gli3 (ובכך מסיר את השפעתו המעכבת על ביטוי הגנים), והקישור גם גורם לאקטיבציה של Gli2, מה שמקדם עוד יותר את ביטוי גני המטרה.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 928-935
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 46
למצע של תרבית תאים הוסיפו TNF-אלפא ולאחר מכן מדדו את רמת הפעילות של NFkB (ע״י מדידת ריכוזו בגרעין התא), את השעתוק של גן A ואת השעתוק של גן B.
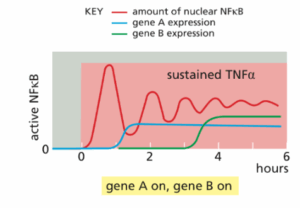
מי מההיגדים הבאים הכי סביר בהתחשב בתוצאות המוצגות בגרף?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ג׳. חלבוני NFkB יוצרים מגוון של הומודימרים והטרודימרים שכל אחד מהם מפעיל את סט הגנים האופייני לו. חלבונים מעכבים הנקראים IkB נקשרים בחוזקה לדימרים הללו ומחזיקים אותם במצב לא-פעיל בציטוזול כל עוד התאים לא עברו סטימולציה.
כאשר TNF-אלפא נקשר לרצפטור שלו, מסלול האיתות מוביל לזרחון של IkB, מה שמסמן את IkB ליוביקוויטילציה ודגרגציה בפרוטאזומים. כעת, NFkB המשוחרר עובר לגרעין ומעורר את שעתוק גני המטרה שלו. בין הגנים המופעלים ע״י NFkB נמצא הגן המקודד לחלבון המעכב IkB. הפעלה זו מובילה לסינתזה מוגברת של IkB אשר נקשר ל- NFkB ומשבית אותו, מה שיוצר לולאת משוב שלילי בדיליי.
המשוב השלילי מייצר שני סוגים של תגובות NFkB, בהתאם למשך הגירוי של TNF-אלפא:
חשיפה קצרה לליגנד מייצרת פולס בודד וקצר של הפעלת NFkB, כי המשוב השלילי מכבה את פעילותו ואין עוד נוכחות של ליגנד כך ש- NFkB לא יופעל שוב.
לעומת זאת, במצב של חשיפה ממושכת ל- TNF-אלפא — החלבון IkB שזה עתה סונתז יעבור יוביקוויטילציה ודגרגציה, כך ששוב פעם ישתחרר NFkB פעיל אשר יוכל לחזור לגרעין ולהפעיל שעתוק. זה למה רואים תנודתיות במסלולים הכוללים משוב שלילי בדיליי (כל עוד הגירוי נמשך).
לפי הגרף המוצג בשאלה, ניתן להבין ששני סוגי התגובות של NFkB מעוררים דפוסים שונים של ביטוי גנים.
ראו את הגרפים למטה:
(A) חשיפה קצרה לליגנד שמייצרת פולס בודד של פעילות NFkB – מעוררת את השעתוק של גן A (ניתן להסיק זאת גם מהגרף המוצג בשאלה מכיוון שרואים שגן A מתבטא בתא כבר מהגל הראשון של NFkB), בעוד שהשעתוק של גן B כלל לא מופעל.
(B) בחשיפה מתמשכת לליגנד שמייצרת פעילות תנודתית ומתמשכת של NFkB – כמובן שהשעתוק של גן A יופעל עוד בהתחלה, אך בהמשך גם השעתוק של גן B מופעל, מה שמצביע על כך שהשעתוק של גן B דורש הפעלה ממושכת של NFkB (מסיבות שאינן מובנות).
שלילת מסיחים:
מסיח א — השעתוק של גן B דורש חשיפה ממושכת לליגנד כפי שהוסבר.
מסיחים ב + ד — לפי הגרף, התנודתיות היחידה היא ברמת הפעילות של NFkB ולא ברמת שעתוק הגנים, פשוט השעתוק של גן A דורש כולה פולס קצר ובודד של הפעלת NFkB, ואילו השעתוק של B דורש הפעלה ממושכת יותר, כלומר כמה פולסים של הפעלת NFkB (ההפעלה הממושכת של NFkB תהיה בתנודות עקב המשוב השלילי).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 934-935
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 47
מי מהסיגנלים החוץ-תאיים הבאים מבקר את שעתוק הגנים בגרעין באופן הישיר ביותר?
במה עוסקת השאלה: איתות תאי, מסלולי איתות אלטרנטיבים בבקרת ביטוי גנים (מצגת ביוכימיה 3 חלק ב׳)
תשובה ד’. הורמונים סטרואידים שייכים למחלקה של סיגנלים חוץ-תאיים שמסוגלים לחצות את הממברנה של תא המטרה, להיקשר לרצפטור החלבוני התוך-תאי שלהם ובכך לשנות את היכולת שלו לשלוט בשעתוק של גנים ספציפיים. בדרך זו, הסיגנלים הללו מבקרים בקרי שעתוק בתוך התא בצורה הישירה ביותר האפשרית. בעצם הרצפטורים של הליגנדים הקטנים וההידרופוביים הללו הם רצפטורים גרעיניים, שהם בקרי שעתוק המְווּסָּתִים ע״י הליגנד שלהם שנכנס לתא ונקשר אליהם.
שלושת המסיחים האחרים מתייחסים לליגנדים של רצפטורי cell-surface, ולכן כדי להשפיע על ביטוי הגנים בגרעין יש ליצור מסלול איתות מורכב שיגיע מהרצפטור בפני שטח התא אל תוך הגרעין.
שלילת מסיחים:
מסיח א — אינסולין נקשר ל- RTK, שהוא רצפטור טרנסממברנלי ולא תוך-תאי כמובן. הרצפטור מזרחן את עצמו וחלבונים נוספים כדי ליצור אתרי עגינה של פוספוטירוזינים על-מנת לבנות קומפלקס איתות שיפעיל את Ras, ואז הוא בתורו יפעיל את קסקדת ה- MAPK ורק שהקינאז האחרון בקסקדה יופעל (Erk), הוא יכנס לגרעין ויזרחן מגוון פקטורי שעתוק ובכך ישפיע על פעילותם.
מסיח ב — גם הליגנד המיטוגני EGF נקשר ל- RTK ובאופן די דומה לאינסולין, הוא משפיע על השעתוק ע״י הפעלת קסקדת Ras-MAPK, שזו כמובן לא דרך ישירה להשפיע על ביטוי הגנים בגרעין.
מסיח ג — רצפטורי ציטוקינים (רצפטורים הקשורים לטירוזין-קינאז) שמפעילים את מסלול JAK-STAT אמנם יוצרים מסלול ישיר יותר לגרעין בהשוואה ל- RTKs ו- GPCRs רבים, אך כמובן שליגנדים של רצפטורים גרעיניים מנצחים בתחרות של מי מבקר את השעתוק באופן ישיר יותר. מסלול האיתות JAK-STAT נחשב לנתיב ישיר יחסית לגרעין בהשוואה לשאר רצפטורי ה- cell-surface מכיוון שלאחר קשירת הליגנד, הטירוזין-קינאז JAK שקשור לרצפטור מגייס ומזרחן ישירות בקרי שעתוק מסוג STATs שנכנסים במהירות לגרעין (אין פה מסלול מורכב שכולל קסקדות של כמה קינאזות או שליחים שניוניים וכו׳).
להרחבה – Alberts, מהדורה שביעית, עמ׳ 935-937
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מסלולי איתות אלטרנטיביים בבקרת גנים + איתות בצמחים
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 48
מי מההיגדים הבאים שגוי בנוגע לאינוזיטול-פוספוליפידים?
במה עוסקת השאלה: איתות תאי
תשובה ה’. פוספוליפידים של אינוזיטול, ובקיצור – פוספטידילאינוזיטול (PI) הוא סוג של פוספוליפיד ממברנלי יוצא דופן מכיוון שהוא יכול לעבור זרחון הפיך במספר אתרים בקבוצת ראש האינוזיטול שלו, כך שנוצרים מגוון של PI מזורחנים הנקראים פוספואינוזיטידים (PIPs).
ה- PIPs מעורבים במגוון מסלולי איתות, ובהקשר זה יש שני אנזימים חשובים שהסובסטרט שלהם זה PIPs מסוימים:
האנזים PI3K מזרחן PIPs בעמדה מספר 3 בראש האינוזיטול. ה- PIP החשוב ביותר שמזורחן בדרך זו הוא PI(4,5)P2, מה שיוצר את PI(3,4,5)P3 שמהווה אתר עגינה למגוון חלבוני איתות שמכילים דומיין PH מתאים, כמו למשל הסרין/תראונין קינאז Akt (שנקרא PKB בלנינג׳ר) מה שתורם לשפעול שלו.
האנזים PLC מזהה את PI(4,5)P2 בלבד ומבקע אותו ליצירת שני שליחים שניוניים – IP3 ו- DAG. לאחר מכן IP3 מדפייז אל ממברנת ה- ER וגורם לשחרור של יוני סידן (שגם נחשב לשליח שניוני).
מסיח א הוא שגוי כי PI(3,4,5)P3 מעורב בשפעול של PKB ואילו יוני סידן ו- DAG, שהוא תוצר הביקוע של PI(4,5)P2, מעורבים בשפעול של PKC.
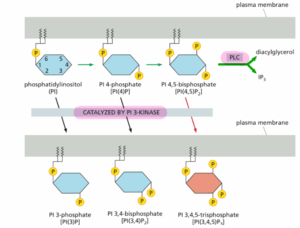
שלילת מסיחים:
מסיח א — נכון. PTEN הוא סוג מסוים של PIPs-פוספטאז. הוא מסיר את הזרחון בעמדה 3 של PI(3,4,5)P3 ובכך מחזיר אותו ל- PI(4,5)P2 ומפסיק את האיתות שמתווך PI3K. כדי לדעת שמוטציות ב- PTEN נמצאות בסוגי סרטן רבים: כאשר PTEN מוטנט זה מוביל להארכת האיתות של PI3K, מה שמקדם גדילה תאית בלתי-מבוקרת.
מסיח ב — נכון. PLC יכול לבקע את PI(4,5)P2 אך לא את PI(3,4,5)P3.
מסיח ג — נכון. PI(3,4,5)P3 הוא תוצר הזרחון של PI(4,5)P2 ולא הסובסטרט של PI3K.
שימו לב ש- PI(4,5)P2 יכול להיות הסובסטרט גם של PLC וגם של PI3K, פשוט התוצר של כל אנזים יהיה שונה וינותב למסלול איתות אחר.
מסיח ד — נכון. PLC יכול לבקע אותו ליצירת IP3 ו- DAG ואז IP3 יגרום לשחרור יוני סידן מה- ER, כך שסה״כ נראה עלייה בריכוז של שלושת השליחים השניוניים הללו.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 920-921
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – קולטני RTK
ספר – Alberts 7th edition
פרק – 15
ביוכימיה 4 שאלה 49
חלבון ב׳ מכיל חומצת אמינו טירוזין שמחוברת לה קבוצת פוספט. איזה interaction domain צריך להיות לחלבון א׳ כדי שהיה לו סיכוי להיקשר לחלבון ב׳?
במה עוסקת השאלה: איתות תאי
תשובה ב’. חלבוני איתות תוך-תאיים רבים מכילים interaction domains. כל סוג של דומיין אינטראקציה נקשר למוטיב מבני מסוים בחלבון או בליפיד. דומייני אינטראקציה מאפשרים לחלבוני איתות להיקשר זה לזה במספר שילובים ספציפיים ובכך ליצור קומפלקסי איתות מורכבים.
יש להכיר ארבעה דומייני אינטראקציה חשובים:
דומיינים מסוג 2 Src homology (דומיין SH2) ודומיינים מסוג phosphotyrosine-binding (דומיין PTB), נקשרים לטירוזינים מזורחנים (פוספוטירוזין) ברצף פפטידי מסוים על גבי רצפטורים פעילים או חלבוני איתות תוך-תאיים. על כן מסיח ב הוא הנכון.
דומיינים מסוג 3 Src homology (דומיין SH3) נקשרים לרצפי חומצות אמינו קצרים ועשירים בפרולין.
כמה דומיינים מסוג pleckstrin homology (דומיין PH) נקשרים לקבוצות ראש טעונות של פוספואינוזיטידים ספציפיים המיוצרים בממברנה הפלזמטית בתגובה לסיגנל חוץ-תאי. הדומיינים הללו מאפשרים לחלבון שהם חלק ממנו לעגון בממברנה ולקיים אינטראקציה עם חלבוני איתות.
דומיינים דמויי-EGF לא קשורים לנושא והמסיח סתם נועד לבלבל. הם מוזכרים בהקשר של דלתא ו- Notch, שמקיימים אינטראקציה זה עם זה דרך הדומיינים דמויי-EGF החוזרניים שלהם.
להרחבה – Alberts, מהדורה שביעית, עמ׳ 883-884
תיוג שאלה:
נושא – ביוכימיה
תת-נושא – חלבונים קושרי-חמצן ואיתות תאי
תת-תת נושא – מבוא לאיתות תאי
ספר – Alberts 7th edition
פרק – 15